Кодирование сигнала в латеральном коленчатом теле и первичной зрительной коре
тормозными и возбуждающими зонами. Таким образом, подобно ганглиозным клеткам сетчатки, для нейронов ЛКТ оптимальным стимулом является контраст, однако они реагируют еще слабее на общее освещение. Изучение рецептивных полей нейронов ЛКТ еще не завершено. Например, в ЛКТ найдены нейроны, вклад которых в работу ЛКТ не был установлен, а также пути, идущие от коры вниз к ЛКТ. Корковая обратная связь необходима для синхронизированной активности нейронов ЛКТ.
Функциональные слои ЛКТ
Почему в ЛКТ на каждый глаз приходится больше одного слоя? Сейчас обнаружено, что нейроны в разных слоях имеют различные функциональные свойства. Например, клетки, находящиеся в четвертых дорзальных мелкоклеточных слоях ЛКТ обезьяны, подобно Ρ ганглиозным клеткам, способны отвечать на свет разных цветов, показывая хорошую цветовую дискриминацию. И наоборот, слои 1 и 2 (крупноклеточные слои) содержат М-подобные клетки, которые дают быстрые («живые») ответы и нечувствительны к цвету, в то время как К слои получают сигналы от "blue-on" ганглиозных клеток сетчатки и могут играть особую роль в цветном зрении. У кошек X и Y волокна (см. раздел «Классификация ганглиозных клеток» заканчиваются в различных подслоях А, С и А1, поэтому специфическая инактивация слоя А, но не С, резко снижает точность глазодвижений. Клетки с "on"- и "off"-центром также подразделяются на различные слои в ЛКТ норки и хорька, и, в некоторой степени, у обезьян. Резюмируя вышесказанное, можно сказать, что ЛКТ является перевалочной станцией, в которой аксоны ганглиозных клеток сортируются таким образом, что соседние клетки получают сигналы от одинаковых регионов зрительных полей, и нейроны, перерабатывающие информацию, организованы в виде кластеров. Таким образом, в ЛКТ очевидной является анатомическая база для параллельной переработки (parallel processing) зрительной информации.
Цитоархитектоника зрительной коры
Зрительная информация поступает в кору и ЛКТ через оптическую радиацию. У обезьян оптическая радиация заканчивается на складчатой пластинке, толщиной около 2 мм (рис. 4). Этот регион мозга — известный как первичная зрительная кора, зрительная зона 1 или V1 — также называется полосатой корой, или «зоной 17». Более старая терминология базировалась на анатомических критериях, разработанных еще в начале XX века. V1 лежит сзади, в области затылочной доли, и может быть распознана при поперечном разрезе по своему особому внешнему виду. Пучки волокон в этой области формируют полоску, ясно видную невооруженным глазом (поэтому зона и называется «полосатой», рис. 4В). Соседние зоны вне зоны полосатости также связаны со зрением. Зона, непосредственно окружающая зону V, называется зоной V2 (или «зона 18») и получает сигналы из зоны V, (см. рис. 4С). Четкие границы так называемой экстрастриарноq зрительной коры (V2-V5) нельзя установить при помощи визуального исследования мозга, хотя для этого разработан ряд критериев. Например, в V2 полосатая исчерченность исчезает, большие клетки расположены поверхностно, и грубые, косо расположенные миелиновые волокна видны в более глубоких слоях
Каждая зона имеет собственное представление зрительного поля сетчатки, спроецированное строго определенным, ретинотопическим образом. Карты проекций были составлены еще в эпоху, когда не было возможно проводить анализ активности отдельных клеток. Поэтому для картирования использовалось освещение пучками света небольших участков сетчатки и регистрация активности коры при помощи большого электрода. Эти карты, а также их современные аналоги, составленные недавно при помощи методов визуализации головного мозга, таких как позитронно-эмиссионная томография и функциональный ядерно-магнитный резонанс, показали, что площадь коры, отведенная на представление центральной ямки, гораздо больше по размерам, чем площадь, отведенная на всю остальную сетчатку. Эти находки, в принципе, соответствовали ожиданиям, поскольку распознавание образов корой осуществляется в основном за счет переработки информации от плотно расположенных в зоне ямки фоторецепторов. Такое представление аналогично расширенному представлению руки и лица в области первичной соматосенсорной коры. Ямка сетчатки проецируется в затылочный полюс коры больших полушарий. Карта периферии сетчатки распространяется в переднем направлении вдоль медиальной поверхности затылочной доли (рис. 5). Из-за перевернутой картины, образуемой на сетчатке при помощи хрусталика, верхнее зрительное поле проецируется на нижнюю область сетчатки и передается в область V1, расположенную ниже шпорной борозды; нижнее зрительное поле проецируется над шпорной бороздой.
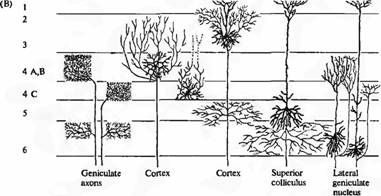
На срезах коры нейроны могут быть классифицированы по их форме. Две основные группы нейронов образуют звездчатые и пирамидные клетки. Примеры этих клеток показаны на рис. 6В. Основные различия между ними заключаются в длине аксонов и в форме тел клеток. Аксоны пирамидных клеток длиннее, спускаются в белое вещество, покидая кору; отростки же звездчатых клеток заканчиваются в ближайших зонах. Эти две группы клеток могут иметь и другие различия, такие как наличие или отсутствие шипиков на дендритах, которые обеспечивают их функциональные свойства. Есть и другие, причудливо названные нейроны (двухбукетные клетки, клетки-люстры, корзинчатые клетки, клетки-полумесяцы), а также клетки нейроглии. Их характерной особенностью является то, что отростки этих клеток направляются в основном в радиальном направлении: вверх и вниз через толщу коры (под соответствующим утлом к поверхности). И наоборот, многие (но не все) их латеральные отростки короткие. Соединения между первичной зрительной корой и корой высшего порядка осуществляется при помощи аксонов, которые проходят в виде пучков через белое вещество, находящееся под клеточными слоями
|
|
|
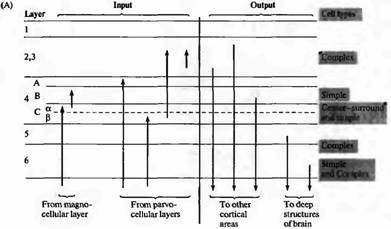
Рис. 7. Связи зрительной коры. (А) Слои клеток с различными входящими и исходящими отростками. Отметим, что исходные отростки из ЛКТ в основном прерываются в 4 слое. Отростки из ЛКТ, идущие от крупноклеточных слоев, преимущественно прерываются в 4С и 4В слоях, в то время как отростки от мелкоклеточных прерываются в 4А и 4С. Простые клетки расположены в основном в слоях 4 и 6, комплексные клетки — в слоях 2, 3, 5 и 6. Клетки слоев 2, 3 и 4В посылают аксоны в другие корковые зоны; клетки в слоях 5 и 6 посылают аксоны к верхнему холмику и ЛКТ. (В) Типичная ветвистость аксонов ЛКТ и кортикальных нейронов кошки. Кроме подобных вертикальных связей, многие клетки имеют длинные горизонтальные связи, идущие в пределах одного слоя к удаленным регионам коры. |
Входящие, исходящие пути и послойная организация коры
Основной особенностью коры млекопитающих является то, что клетки здесь расположены в виде 6 слоев в пределах серого вещества (рис. 6А). Слои сильно различаются по внешнему виду, в зависимости от плотности расположения клеток, а также толщины каждой из зон коры. Входящие пути показаны на рис. 7A с левой стороны. Исходя из ЛКТ, волокна в основном заканчиваются в слое 4 с небольшим количеством связей, образуемых также в слое 6. Поверхностные слои получают сигналы из области подушки таламуса (pulvinar zone) или других зон таламуса. Большое количество клеток коры, особенно в области слоя 2, а также в верхних частях слоев 3 и 5 получают сигналы от нейронов, также расположенных в пределах коры. Основная масса волокон, идущих от ЛКТ в слой 4, затем разделяется между различными подслоями.