Кодирование сигнала в латеральном коленчатом теле и первичной зрительной коре
Исходящие из слоев 6, 5, 4, 3 и 2 волокна показаны справа на рис.7А. Клетки, посылаюшие эфферентные сигналы из коры, могут также управлять внутри корковыми соединениями между разными слоями. Например, аксоны клетки из слоя 6, кроме ЛКТ, могут также направляться в один из других кортикальных слоев, в зависимости от типа ответа этой клетки34). На основании подобного строения зрительных путей можно представить следующий путь зрительного сигнала: информация с сетчатки передается на клетки коры (в основном, в слой 4) аксонами клеток ЛКТ; информация передается из слоя в слой, от нейрона к нейрону по всей толщине коры; переработанная информация пересылается в другие зоны коры при помощи волокон, направляющихся вглубь белого вещества и возвращающихся обратно в область коры. Таким образом, радиальная или вертикальная организация коры дает нам основания полагать, что колонки нейронов работают как отдельные вычислительные единицы, обрабатывая различные детали зрительных сцен и пересылая полученную информацию далее в другие регионы коры.
Разделение входящих волокон от ЛКТ в слое 4
Афферентные волокна ЛКТ заканчиваются в слое 4 первичной зрительной коры, который имеет сложную организацию и может быть исследован как физиологически, так и анатомически. Первой особенностью, которую мы хотим продемонстрировать, является разделение входящих волокон, идущих от разных глаз. У взрослых кошек и обезьян клетки в пределах одного слоя ЛКТ, получая сигналы от одного глаза, посылают отростки к строго определенным скоплениям клеток коры в слое 4С, отвечающим именно за этот глаз. Скопления клеток сгруппированы в виде чередующихся полосок или пучков кортикальных клеток, получающих информацию исключительно от правого или левого глаза. В более поверхностно и глубже расположенных слоях нейроны управляются обоими глазами, хотя обычно с преобладанием одного из них. Хьюбель и Визель провели оригинальную демонстрацию разделения информации от разных глаз и преобладания одного из них в первичной зрительной коре при помощи электрофизиологических методов. Они использовали термин «глазодоминантные колонки» (ocular dominance columns) при описании своих наблюдений, придерживаясь концепции кортикальных колонок, разработанной Маунткаслом для соматосенсорной коры. Серия экспериментальных методик была разработана для демонстрации чередующихся групп клеток в слое 4, получающих информацию от правого или левого глаза. Вначале было предложено нанести небольшое повреждение в пределах только одного слоя ЛКТ (напомним, что каждый слой получает информацию только от одного глаза). Если это сделать, то дегенерирующие терминали появляются в слое 4, образуя определенный паттерн чередующихся пятен, которые соответствуют зонам, управляемым глазом, посылающим информацию в поврежденную область ЛКТ. Позднее потрясающая демонстрация существования особого паттерна глазного доминирования была выполнена, используя транспорт радиоактивных аминокислот из одного глаза. Эксперимент состоит в том, что в глаз вводится аминокислота (пролин или лецитин), содержащая атомы радиоактивного трития. Инъекция проводится в стекловидное тело глаза, из которого аминокислота захватывается телами нервных клеток сетчатки и включается в состав белка. Со временем помеченный таким образом белок транспортируется в ганглиозные клетки и по волокнам зрительного нерва в их терминали в пределах ЛКТ. Замечательной особенностью является то, что эта радиоактивная метка также передается от нейрона к нейрону через химические синапсы. В конечном итоге метка попадает в окончания волокон ЛКТ в пределах зрительной коры.
На рис. 8 показано расположение в пределах слоя 4 радиоактивных терминалей, образованных аксонами клеток ЛКТ, связанных с глазом, в который вводилась метка
|
Рис. 8. Глазодоминантные колонки в коре обезьяны, полученные при помощи введения радиоактивного про лина в один глаз. Ауторадиограммы, снятые при темнопольном освещении, где белым показаны зерна серебра. (А) Сверху рисунка срез проходит через слой 4 зрительной коры под углом к поверхности, образуя перпендикулярный срез колонок. В центре слой 4 был срезан горизонтально, показывая, что колонка состоит из удлиненных пластинок. (В) Реконструкция из множества горизонтальных срезов слоя 4С у другой обезьяны, у которой инъекция проводилась в илсилатеральный глаз. (Любой горизонтальный разрез может выявить |
лишь часть слоя 4, что обусловлено кривизной коры.) Как в А, так и в В колонки зрительного доминирования выглядят как полоски равной ширины, получающие информацию либо от одного, либо другого глаза. |
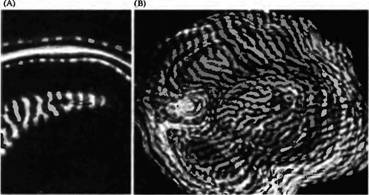
располагают непосредственно над зрительной корой, поэтому такие участки выглядят как белые пятна на темном фоне фотографии). Пятна от меток перемежаются с зонами без меток, которые получают информацию от контралатерального глаза, куда не вводилась метка. Расстояние от центра до центра между пятнами, которые соответствуют глазодоминантным колонкам, составляет приблизительно 1 мм.
На клеточном уровне сходная структура была выявлена в слое 4 при помощи введения пероксидазы хрена в отдельные направляющиеся в кору аксоны нейронов ЛКТ. Аксон, показанный на рис. 9, идет от нейрона ЛКТ с "off"-центром, отвечающим короткими сигналами на тени и движущиеся пятна. Аксон заканчивается в двух различных группах отростков в слое 4. Группы меченых отростков отделены пустой немеченной зоной, соответствующей по своим размерам территории, отвечающей за другой глаз. Подобного рода морфологические исследования расширяют границы и позволяют более глубоко понять оригинальное описание колонок глазного доминирования, составленное Хьюбелем и Визелем в 1962 году.