Риниофиты. Бесполое размножение. Спорангии и спорогенез у древних высших растений
Риниофиты (Rhyniophyta), псилофиты (Psilophyta), самая древняя и примитивная вымершая группа (отдел) высших растений. Характеризовались верхушечным расположением спорангиев и равноспоровостью, отсутствием корней и листьев, дихотомическим или дихоподиальным (псевдомоноподиальным) ветвлением, примитивным анатомическим строением. Проводящая система — типичная протостела. Протоксилема располагалась в центре ксилемы; метаксилема состояла из трахеид с кольчатыми или (реже) лестничными утолщениями. Опорные ткани отсутствовали. Р. ещё не обладали способностью ко вторичному росту (меристемы у них были только верхушечные). Спорангии примитивные, от шаровидных (диаметром около 1 мм) до продолговато-цилиндрических (длиной до 12 мм), толстостенные. Гаметофиты. достоверно не известны (некоторые авторы считают гаметофитами горизонтальные корневищеподобные органы — так называемые ризомоиды). Риниофиты. произрастали на влажных и болотистых местах, а также в прибрежном мелководье. Отдел Риниофиты. включает один класс — риниопсиды (Rhyniopsida) с двумя порядками — Rhyniales (семества Cooksoniaceae, Rhyniaceae, Hedeiaceae) и Psilophytales (семейство Psilophytaceae). Для порядка Rhyniales характерны дихотомическое ветвление и тонкая, слабо развитая стела. Ксилема из трахеид с кольчатыми утолщениями. Древнейший представитель Риниофит. — род куксония, первоначально обнаруженный в Уэльсе в отложениях конца силурийского периода (около 400 млн. лет назад). Наиболее полно изучены нижнедевонские роды — риния и отчасти хорнеофит, у которого ризомоид (вверх от него отходили стебли, вниз — многочисленные ризоиды) был расчленён на чётковидно расположенные клубневидные сегменты, лишён проводящих тканей и целиком состоял из паренхимных клеток. Полагают, что в процессе эволюции ризомоиды. дали начало корням. У обоих родов стенка спорангия была многослойной, покрытой кутикулой. Хорнеофит характеризуется своеобразной спороносной полостью, которая образует купол, сводообразно покрывающий центральную колонку стерильной ткани, представляющей собой продолжение флоэмы стебля. Этим хорнеофит напоминает современный сфагнум. В семейства риниевых включают также род тениокрада, многие виды которого образовывали подводные заросли в среднем и верхнем девоне. В отдельное семейства хедеевых иногда выделяют нижнедевонские роды хедея и яравия. Нижнедевонский род сциадофит, обычно выделяемый в отдельное семейства сциадофитовых, — небольшое растение, состоявшее из розетки простых или слабо дихотомированных тонких стеблей со стелой. Для порядка Psilophytales характерны дихоподиальное ветвление и более сильно развитая стела. У наиболее известного рода — псилофит (из нижнедевонских отложений в Восточной Канаде) — неравно развитые ветви образовывали ложную главную ось дихоподия с более тонкими боковыми ветвями: стебель был окружен кутинизированной эпидермой с устьицами; поверхность стебля была голая или покрыта шипами длиной 2—2,5 мм, концы которых дисковидно расширялись, что, вероятно, указывало на их секреторную роль. Спорангии раскрывались продольной трещиной. К псилофиту близки нижнедевонские роды тримерофит и пертика.
Бесполое размножение. Спорангии
В предыдущей главе мы попытались проследить эволюцию спорангиеносных частей у архегоняальных растений. Теперь перейдем к рассмотрению самого процесса размножения. Попытаемся уловить основные моменты эволюции в развитии спорангиев и спор.
Как известно, у папоротникообразных существует два типа развития спорангия: евспорангиатный и лептоспорангиатный. При развитии спорангия по евепорангиатному типу в образовании его участвует целый участок ткани или во всяком случае группа клеток. Самый же спорангий в большинстве случаев отличается крупными размерами и толстой, многослойной стенкой.
Лептосиорангиатный тип характеризуется тем, что спорангий возникает из одной, эпидермической клетки, имеет небольшую величину и тонкую, однослойную стенку, снабженную более или менее ясно выраженным кольцом. По евепорангиатному типу развиваются спорангии у Psilotales, плаунов, хвощей, ужовников, маратциевых, у всех голосемянных. По этому же типу, вероятно, развивались спорангии у ископаемых представителей Lycopsida и Sphenopsida.
По лептоспорангиагному типу развиваются спорангии у настоящих папоротников. Примером евшорангиатного типа может служить развитие спорангия плауна — L. clavatura (рис. 69). По исследованиям П. А. Баранова (1924) спорангий L. clavatum закладывается на верхней стороне спорофилла, ближе к его основанию. Начало ему дает группа поверхностных клеток, которые уже на очень ранних стадиях несколько отличаются своим содержимым от соседних. Клетки эти делятся стенками, параллельными поверхности спорофилла, на наружные и внутренние. Из внутренних в дальнейшем образуется археопорий, из наружных — стенки спорангия. И те и другие клетки энергично делятся, в результате чего образуется бугорок. В дальнейшем в этом бугорке дифференцируется археспорий и двуслойная стенка спорангия. Клетки археспория отличаются более крупными размерами, крупными ядрами и густой протоплазмой. Между археспорием и стенкой спорангия образуется выстилающий слой — тапетум. Он отделяется от внутреннего слоя стенки спорангия, а в нижней части спорангия дифференцируется из прилежащей к археспорию ткани. Зачаток спорангия растет неравномерно: нижняя часть его отстает от верхней, в из нее образуется ножка спорангия. Клетки археспория обособляются и превращаются в материнские клетки спор, которые обычным путем дают тетрады и опоры.
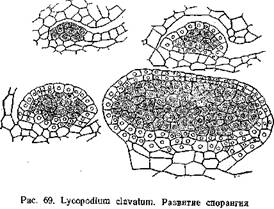
Также в общих чертах развиваются спорангии у селагинелл, хвощей и поилотов. Эволюция спорангиев выразилась в выработке приспособления для раскрывания спорангия и в возникновении особого Слоя, играющего роль в питании развивающихся спор и образовании у них сложных оболочек — тапетума или выстилающего слоя. Задачу раскрывания спорангия принимает на себя Наружный слой его стенки, превращаясь в так называемый экзотеций. Стенки его клеток утолщаются, причем утолщения эти у различных представителен папоротникообразных носят различный характер (балки, спирали и т. д.). Среди утолщенных клеток проходят ряды клеток снеутолщенными клетками (stomium). По ним проходит трещина, разрывающая стенку спорангия.
Выстилающий слой располагается между стенкой спорангия и спорогенным комплексом. Он отчленяется или от стенки спорангия (Lyoopodium) или от самого спорогенного комплекса, то есть наружные Слои этого последнего превращаются в тапетум (Ophioglossum, Equisetum). При этом различают два рода тапетума: секреторный и периплазмодиальный.
У секреторного тапетума стенки клеток сохраняются до конца его существования, и тапетум все время представляет собой слой клеток, облекающий внутреннюю поверхность спорангия. Но нередко клетки тапетума увеличиваются в размерах. При этом изменяется и характер их содержимого. Считают, что клетки секреторного тапетума выделяют теили иные вещества, которые участвуют в образовании спор и покрывающих их оболочек. Однако более подробно роль эта остается невыясненной.