Риниофиты. Бесполое размножение. Спорангии и спорогенез у древних высших растений
Евопорангиатный тип спорангиев встречается у наиболее древних и примитивных групп среди ныне живущих папоротникообразных. Он же характерен) и для ископаемых папоротникообразных. Это заставляет нас смотреть на евшорангий как на древний, примитивный тип спорангия, от которого уже произошел лептосггорангий, свойственный к тому же более молодым группам папоротников (Filices). Об этом говорит и то, что, как показывают сравнительные исследования, между евопорангиатный я лептоспорангиатным типами развития существует ряд постепенных переходов, связывающих их между собой. Вместе с тем эти переходы указывают на ход эволюции лептоспорангия.
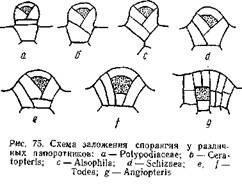
На рис. 75 дана схема заложения опорангия у различных папоротников: рис. а относится к типичному лаптоспорангию (сем. Polypodiaeeae), а рис. g— к типичному евспорангию (Angiopteris). Между ними располагаются промежуточные типы, принадлежащие к различным родам. Понятно, эволюция лептоспорангия совершалась в направлении, обратном расположению рисунков. У Angiopteris археспорий закладывается глубоко под поверхностью и самая археспориальная клетка имеет форму куба: ограничивающие ее стенки пересекаются под прямым углом. У Todea (сем. Osmundaceae) наблюдаются два варианта развития опорангия. В одном случае ([) археспорий имеет в разрезе трапециевидную форму, благодаря тому, что антиклины, ограничивающие археспорий, наклонены под некоторым углом друг к другу. В другом случае (более мелкие спорангии) эти антиклины пересекаются, и археспорий принимает форму четырехгранной пирамиды, обращенной вершиной вниз, то есть форму, характерную для археспория лептоепорангия. И у Angiopteris и у Todea в образовании спорангия принимают участие и прилежащие к археспорию клетки. Развитие спорангия у Schiziaea (d), Alsophila(с) и Ceratopteris (Ь) представляет переход к типичному лептоспорангиатному типу (а). Археспорий окончательно принимает форму четырехгранной пирамиды и все меньшее число эпидермических клеток идет на развитие спорангиев. У Polypo-diaceae (а) весь спорангий развивается из одной эпидермической клетки.
Таким образом, эволюция спорангия у папоротников шла в направлении редукции, которая и приведет к образованию лептоспорангия. Эволюция последнего, кроме того, выразилась в создании длинной и тонкой ножки, выработке кольца и в общем уменьшении количества производимых спорангием спор. В результате получился типичный лептоспорангий малой величины с длинной ножкой и однорядным кольцом. Число спор, образующихся в спорангии, невелико, но число спорангиев сильно возрастает, благодаря чему общее количество спор, производимых растением, не только не уменьшается, но и весьма увеличивается.
По характеру спор среди археголиальных мы различаем две труппы растений — равноепоровые и разноюлоровые (гомо- и гегероспоровые) формы. И те и другие встречаются во всех классах папоротникообразных; голосемянные—исключительно тетероспоровые. Это показывает, что гетероспория возникала в разных типах и классах самостоятельно я независимо. Естественно встает вопрос, какой тип спор мы должны считать первоначальным, исходным, иначе, развилась ли гетероспория из гомоспории или, наоборот, гетероспория дала начало гомоспории. Прямых указаний на то или иное решение вопроса мы не имеем, теоретически же возможно допустить и тот и другой ход эволюции. Однако, исходя из того факта, что гетероспория всегда связана с сильной редукцией гвметофита, в принимая во внимание то, что примитивный гаметофит должен был быть хорошо развитым образованием, правильнее будет предположить, что гомоопория является примитивным признаком и первоначально все опоры были гомоспорами, то есть при прорастании давали обоеполые заростки, несущие и антеридии и архегонии. В дальнейшей эволюции произошла дифферевцировка их на микроспоры и мегаспоры — мужские и женские опоры. Указание на такого рода дифференцирозку мы находим и среди мхов, согласно исследованиям сравнительно недавнего времени.
У Utota bruchii, например, величина спор, образующихся в одном и том же спорангии, сильно колеблется (от 21 до 32 (х). Однако связана ли эта разница в величине с половой дифференцировкой, остается невыясненным. Но для форм, близких к Ulota bruchii, из того же семейства Orthotriehaeeiae, именно для Macromitrium blumeivH Schlotheimia konigsbergii таковая дифференциревка была доказана М. Ф. Фдейшером. Споры у этик мхов также сильно разнятся в размера и, как показал М. Флейшер, яри ирорастании их из более крупных спор вырастают всегда карликовые мужские растения. Из мелких же спор развиваются нормальные женские особи.
М. Флейшер нашел также карликовые мужские и женские растения у Trismegistia brauniana (из Hypnobryales), хотя споры у этого мха внешне не отличаются друг от друга по размерам. Внутренняя половая дифференцировка, таким образом, у них имеется. Карликовые мужские растения Флейшер нашел и у многих других мхов.
Позднее гетероспория была обнаружена М. Эрнет-Шварценбах (Ernst-Schwarzenbach,1939) у ряда видов Macromitrium. Этим автором было исследовано 35 видов p. Macromitrium. Из них 16 видов оказались гомостюровыми, 14— явно гетероспоровыми, 5—неясно гетероспоровыми. Например у salakense микроспоры имеют в- среднем 19,6 ц в диаметре, мегаспоры — 36,7 ц. Микроспоры имеют желтую окраску и одеты гладкой оболочкой, мегаспоры — зеленые, оболочка их — бородавчатая. Отношение тех и других— 1 : 1. На выросшей из микроспор протонеме через 3—4 месяца после посева образовались карликовые растения с антеридиями. На протонеме, развившейся ив мегаспор, появились Стебля, но к моменту окончания опытов половых органов на них еще не было. На них, по-видимому, при дальнейшем развитии -должны были появиться архегонии.
Как видно из сказанного, между данными М. Флейшера и М. Эрнст-Шварценбах имеется известное противоречие. По М. Флейшеру, микроспоры являются женскими спорами, мегаспоры — мужскими, по М. Эрнст-Шварценбах, наоборот, то есть мхи ведут себя в этом отношении так же, как папоротникообразные. Вопрос о гегероспории у мхов требует, следовательно, дальнейшего исследования. Однако и в настоящий момент мы можем констатировать, что у лиственных мхов гетероспория в той или иной форме существует. Указаний на гетероепорию у печеночников нет.
Большинство гомоепоровых папоротникообразных обладает обоеполыми заростками. Это указывает на то, что в спорах их нет половой дифференцировки и все споры и внешне и внутренне вполне равноценны. Однако зачатки гегероспории мы можем уловить и у них. Так, по свидетельству К. Гебеля, споры типичного папоротника Dennstedtia pimctilobula при посеве дают однополые заростки, то есть часть их развивается в мужские, часть в женские заростки. То же самое наблюдается и у некоторых хвощей, как например, у Equisetum limosum, E. hiemale, E. silvestre. Споры стих. хвощей дают мужские и женские заростка, отличающиеся друг от друга размерами, строением и даже окраской. Мужские заростки — меньшей величины, более - простого строения по сравнению с женскими и более светлой, желтоватой окраски. И у Dennstedtia, и у названных выше хвощей пол споры, возможно, определяется генотипически, так что если все споры -по величине и форме одинаковы, то внутреннее их различие — весьма резкое. У других хвощей, как у Е. debile, E. laevigatum или Е. arvense, заростки — обоеполые, причем архегонии на них развиваются раньше антеридиев. Но наряду с обоеполыми у этих хвощей образуются и чисто мужские заростки; количество их весьма зависит от тех условий, при которых прорастают споры и развиваются заростки.