Совершенствование полуэмпирических методов рационального использования биологических ресурсов водоемов
Один из важных способов определения оптимальной интенсивности вылова и селективности лова связан с применением для этой цели модификаций уравнения Баранова-Бивертона-Холта. Критерием оптимальности при обосновании этих показателей служит величина улова на единицу пополнения промыслового стада. Для оценки оптимальных значений показателей интенсивности и селективности рыболовства целесообразно использовать модификацию этого уравнения, предложенную А.В. Мельниковым (1987), которое описано в гл. 5 в связи с использованием этого уравнения для оценки запасов.
В главе 5 рассмотрены также особенности определения оптимального значения интенсивности вылова и селективности лова с применением конечно- разностных уравнений при переменном пополнении, росте и естественной смертности рыб.
4.3. Определение оптимальной интенсивности вылова и селективности лова с учетом эффективности использования биомассы поколения
Из рассматриваемого условия рыбу следует вылавливать в возрасте, соответствующем кульминации ихтиомассы поколения. Выловить всю рыбу в этом возрасте практически невозможно. Чтобы повысить эффективность использования ихтиомассы, лов рыбы начинают в возрасте, меньшем возраста tм кульминации ихтиомассы, а заканчивают позже этого возраста. Чем меньше интенсивность вылова, тем раньше необходимо начинать лов рыбы данного поколения из рассматриваемого условия. Hо возраст, в котором начинают облавливать рыбу, зависит в основном от размера ячеи. Также от размера ячеи зависит и степень использования ихтиомассы поколений рыб. Вот почему одним из показателей при обосновании мер регулирования рыболовства служит коэффициент использования биомассы поколения
кб = Q / Qм, (4.1)
где Q - масса улова поколения при лове заданной интенсивности и заданном размере ячеи; Qм - масса поколения в возрасте кульминации ихтиомассы.
Регулируя интенсивность вылова, размер ячеи, а иногда также промысловую меру на рыбу и допустимый прилов рыб непромысловых размеров, можно добиться повышения коэффициента кб.
Для установления зависимости коэффициента использования биомассы поколения кб от различных факторов определим сначала Qм и Q, входящие в (4.1).
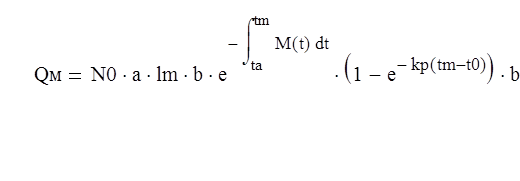 Масса улова Qм равна биомассе поколения в возрасте tм, соответствующем кульминации ихтиомассы с учетом естественной смертности рыб:
Масса улова Qм равна биомассе поколения в возрасте tм, соответствующем кульминации ихтиомассы с учетом естественной смертности рыб:
где Nо - численность поколения в возрасте tа, когда рыба становится объектом лова; M(t) - функция, характеризующая изменение коэффициента естественной смертности с возрастом; а и b - коэффициенты в уравнении масса-длина; lм, кр и tо - параметры уравнения Берталанфи.
Возраст tм находят различными способами. Если в выражении (4.2) tм считать текущим временем t, то можно построить график изменения относительной биомассы с возрастом, полагая Nо= 1. Вершине этого графика соответствует tм.
Если мгновенный коэффициент естественной смертности М считать постоянным, а рост рыбы принимать в соответствии с уравнением Берталанфи, то возраст tм можно определить по формуле Катти (Рикер, 1979)

Для определения возраста tм удобно воспользоваться методикойП.В. Тюрина (1962), суммируя массы отдельных возрастных групп с учетом годовой убыли от естественной смертности и строя график изменения массы поколения, по которому определяют tм.
Биомасса улова
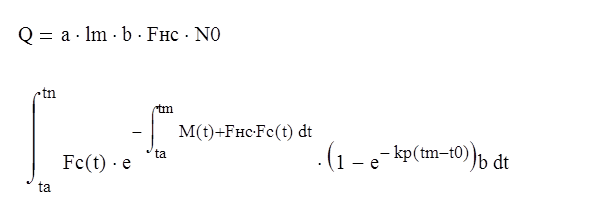 |
где tп - предельный возраст рыб в уловах; Fнс - коэффициент промысловой смертности при условно неселективном лове,т.е. когда функция кривой селективности Fс (t) = S(t) =1.
Подставляем Qм и Q в формулу (9.1):
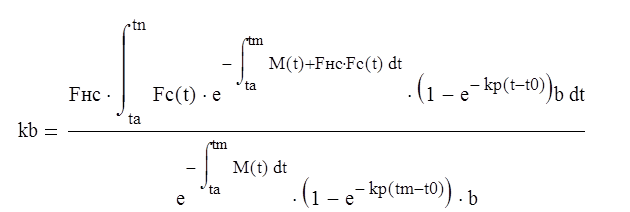
,(4.5)
По формуле (4.5) кб определяют для различной интенсивности вылова, размера ячеи и других влияющих факторов. В частности,одному из размеров ячеи соответствует максимальное значение коэффициента, и при таком размере ячеи ихтиомасса поколения используется наиболее полно. Этот размер ячеи сравнивают с размерами ячеи, полученными из других соображений, и выбирают компромиссный вариант.
В общем случае исследуют зависимость коэффициента использования биомассы поколения одновременно от размера ячеи (селективности лова) и от коэффициента промысловой смертности при условно неселективном лове Fнс (интенсивности лова) и определяют, при каких практически возможных значениях селективности лова и интенсивности промысла биомасса поколения используется наиболее рационально.
В соответствии с рассмотренной методикой при оценке кб учитывают всех рыб в улове и биомассу всех облавливаемых рыб. Кроме того, представляет интерес случай, когда в улове учитывают только рыб промысловых размеров, а при оценке максимальной биомассы- всех рыб или только рыб промысловых размеров.
Выражения для коэффициента использования биомассы поколения для рассмотренных случаев несложно получить из (4.5). Важно, что с учетом дополнительных условий величина кб зависит от промысловой меры на рыбу и допустимого прилова рыб непромысловых размеров и их также можно оптимизировать с учетом рассматриваемого критерия оптимальности. При определении коэффициента использования биомассы в этих случаях учитывают, что возраст кульминации ихтиомассы иногда меньше возраста, соответствующего промысловой мере на рыбу.
Для понимания сущности оценки допустимого значения кб и оценки регламентирующих интенсивность и селективность лова показателей иногда полезно представить его в виде произведения двух коэффициентов кб' и кл. Первый из них характеризует эффективность использования биомассы поколения при неселективном лове, а второй учитывает влияние селективности лова.
Коэффициент кб' получают из (4.5), считая Fc (t)= S(t) = 1:
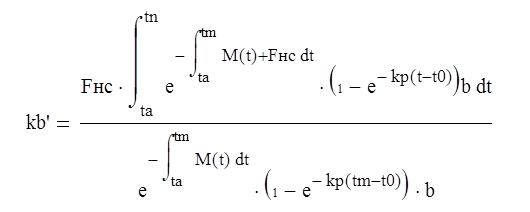
(3.6)
Отличие коэффициента кб' от единицы учитывает, что рыбу вылавливают в возрасте, не соответствующем кульминации ихтиомассы с учетом ограниченной интенсивности вылова. Величина коэффициента кб' является предельно возможным значением коэффициента кб при заданных темпах роста, промысловой смертности и интенсивности вылова.
Коэффициент влияния селективности орудия лова
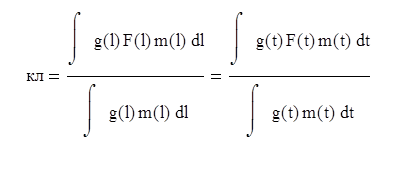
Рассчитав значения коэффициента кл для различного размера ячеи, можно построить график кл = f(А ). Коэффициент кл учитывает потери рыбы, обусловленные уходом рыбы через ячею, и служит самостоятельным показателем при обосновании мер регулирования рыболовства. Величина этого коэффициента при реальных размерах ячеи обычно не меньше 0,7-0,75.
Определение коэффициентов кб, кб' и кл важно не только в связи с обоснованием показателей, регламентирующих интенсивность и селективность рыболовства, но имеет и самостоятельное значение, в частности, как показателей, характеризующих механизм и особенности эксплуатации запасов промысловых рыб.