Совершенствование полуэмпирических методов рационального использования биологических ресурсов водоемов
Биомеханический принцип селективности обусловлен различной плавательной способностью рыб, особенностями их поведения в потоке воды. Влияние этих факторов сказывается на селективности всех подвижных сетных орудий лова и рыбонасосных установок. Селективность с использованием биомеханического принципа регулируют в основном выбором скорости перемещения сетных орудий лова, скорости всасывания и показателей перемещения залавливающего устройства рыбонасосных установок.
Биофизический принцип селективности учитывает неодинаковую реакцию рыб разного вида, размера и пола на действие физических полей естественного происхождения, орудий лова и средств интенсификации лова.
Геометрический принцип селективности связан с размерами и формой зоны облова орудия лова, особенностями распределения рыбы в зоне облова по виду, размеру и полу.
Биологический принцип селективности орудий лова вытекает из неодинаковой способности рыб разного вида, размера и пола избегать орудия лова и уходить из зоны облова, прежде всего, благодаря различному врожденному и приобретенному опыту.
Геометрический и биологический принципы селективности с трудом поддаются количественной оценке, имеют меньшее значение, по сравнению с другими видами селективности, и в нашей работе не рассматриваются.
Далеко не всегда можно разделить эффект селективности, основанный на том или ином принципе, т.к. часто их проявления взаимосвязаны. Так, показатели биомеханической, геометрической и биологической селективности во многом зависят от действия на рыбу физических полей; механическая селективность в некоторой степени зависит от действия тех же полей, показателей перемещения рыбы и орудия лова (Судаков, 1998).
Наиболее часто различают пространственную, временную и пространственно-временную селективности промысла в зависимости от размеров промысловой акватории и промыслового времени при оценке селективности промысла. Практически обычно рассматривают пространственно-временную селективность, имея в виду одновременно ограничения промысла в пространстве и времени (Мельников, 1999).
Также рассматривают пространственную, временную и пространственно- временную селективность рыболовства, учитывая работу орудий лова и распределение промысла в определенных пространственных и временных границах.
Кроме рассмотренного вида селективности промысла, которая зависит исключительно от распределения объекта лова и промысловых единиц в районе промысла и которую можно назвать пространственно-временной селективностью промысла, иногда определяют статистическую селективность промысла, которая характеризуется усредненным показателем селективности лова отдельных промысловых единиц и которая зависит также от селективных свойств отдельных орудий (способов) лова.
Иногда селективность промысла сложно отделить от селективности орудий лова. Например, селективность орудий лова, основанная на геометрическом принципе, как и селективность промысла, зависит от особенностей распределения рыбы в водоеме.
В современном рыболовстве наибольшее значение имеет механический принцип селективности, связанный, прежде всего, с селективными свойствами ячеи.
При лове подвижными орудиями лова иногда важное значение имеет биомеханическая селективность; при использовании физических полей как средства интенсификации лова - биофизическая селективность, при высокой неравномерности распределения рыбы в зоне облова - геометрическая селективность орудий лова.
1.2. Показатели селективности лова при отцеживании
За основу оценки селективных свойств сетных полотен примем выражения для кривой селективности в виде логистической кривой (Мельников А.В.,1985). Уточнения в этом выражении связаны, прежде всего, с отсутствием для многих видов лова доли рыб, не подверженных селективному действию ячеи, обусловленной большим количеством рыб в улове. С другой стороны, установлено, что и при малом количестве рыб в улове некоторая доля рыб не подвержена селективному действию ячеи, поскольку часть рыбы оказывается у сетного полотна в положении, неудобном для прохода через ячею.
С учетом этих дополнений математическая модель кривой селективности концентрирущих частей отцеживающих орудий лова и ловушек через параметры кривой, коэффициент селективности кs, диапазон селективности Ds, и долю рыб, не подверженных селективному действию ячеи aнc', и размер ячеи А имеет вид:
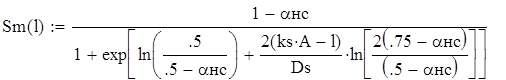
(1.1)
где αнc – доля рыб, не подверженных селективному действию ячеи, при любом количестве рыб в улове; ks – коэффициент селективности; Ds – диапазон селективности; А – внутренний размер ячеи; l – длина рыбы.
Хаpактеp кpивой селективности ячеи и сетного мешка можно уточнить, если диапазон селективности pассматpивать pаздельно в интеpвале от l0,25 до l0,5 и от l0,5 до l0,75 и другими способами(Мельников, 1985, 1991 и т.д.). Однако, как показали дальнейшие исследования, эти уточнения в практических расчетах можно не учитывать. Пример семейства кривых селективности для различного размера ячеи приведен на рис. 1.1.
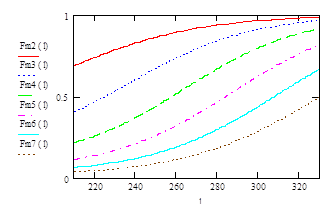
Рис1.1 Семейство кривых селективности для различного размера ячеи.
Долю рыб, не подверженных селективному действию сетного мешка при затеняющем действии улова обычно определяют по формуле (Мельников, 1985):
![]()
(1.2)
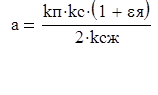
где ; kп – коэффициент полноты тела рыбы; kсж – коэффициент сжатия тела рыбы; kc – коэффициент соответствия рабочей формы ячеи форме поперечного сечения тела рыбы; εя - рабочее относительное удлинение ячеи с внутренним размером А; lо и lн - длина рыб, соответствующая накопленной частоте встречаемости среди попадающих в сетной мешок 0,02-0,05 и 0,13- 0,25.
С учетом небольших уловов при работе ряда орудий лова получено следующее выражение для оценки доли рыб, не подверженных селективному действию ячеи:
![]()
(1.3)
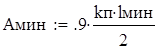
(1.4)
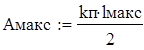
(1.5)
где lмакс и lмин - соответственно максимальный и минимальный размер рыб в облавливаемых скоплениях.
Пример зависимости доли рыб, не подверженных селективному действию сетного мешка, от размера ячеи приведен на рис. 1.2.
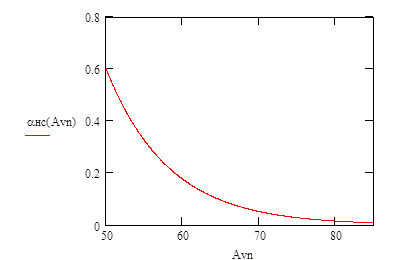
Рис 1.2 Зависимости доли рыб, не подверженных селективному действию сетного мешка, от размера ячеи.
Коэффициент селективности сетного мешка, по А.В. Мельникову (1985),
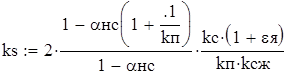
(1.6)