Сравнительная характеристика и морфофизиологические основы мышления животных
Сравнительная характеристика уровня развития элементарного мышления у животных разных таксономических групп. Некоторые показатели уровня развития мозга (относительные размеры его высших отделов, а также сложность нейронного строения и межнейронных связей), которые коррелируют со степенью развития элементарной рассудочной деятельности. Данные о дифференцированном влиянии разрушения одних и тех же структур на рассудочную деятельность и способность к обучению. Показано, что животным, обладающим хорошо развитым мозгом, доступны более сложные по логической структуре и более разнообразные задачи. Характеристики рассудочной деятельности наиболее продвинутых животных, степень их сходства с психикой человека и отличия от психики других животных. Обсуждение параллелизма в развитии рассудочной деятельности представителей разных классов.
Современное исследование любого аспекта поведения только тогда может считаться полноценным, если авторы рассматривают его развитие, механизмы, эволюцию и адаптивное значение, не ограничиваясь наблюдениями и описаниями. В предыдущих главах был рассмотрен практически весь диапазон форм элементарного мышления, доступных животным. Одни формы мышления достаточно примитивны и встречаются у низкоорганизованных животных, другие — более сложные и доступны видам, наиболее продвинутым в эволюционном развитии. Интересно рассмотреть, на каких этапах филогенеза они возникли, как происходило их усложнение в эволюционном ряду, а также выяснить, какие особенности строения мозга обеспечивают проявление того или иного уровня развития рассудочной деятельности.
1. «Сложное обучение» и уровень развития животных.
В многочисленных исследованиях было неоднократно продемонстрировано, что скорость, прочность и «точность» формирования обычных УР (как классических, так и инструментальных) достаточно близки у позвоночных самого разного филогенетического уровня и практически не зависят от сложности строения мозга. Чтобы оценить степень развития более сложных когнитивных функций, необходимо было разработать другие процедуры обучения животных, которые требовали бы улавливания общего «правила», «алгоритма» решения серии однотипных задач. Предполагалось, что их использование могло бы выявить различия в обучении и когнитивных процессах, которые коррелировали бы со сложностью строения мозга.
В лаборатории Л. Г. Воронина в 60-е годы на животных разных таксономических групп (рыбах, пресмыкающихся, птицах и млекопитающих) исследовали скорость формирования «цепных» УР. Оказалось, что «цепи» УР легче вырабатываются и дольше сохраняются у животных с более развитым мозгом.
Методом многократных переделок УР уже в первых работах убедительно показано, что успешность обучения в целом зависит от уровня организации мозга животного, по крайней мере в пределах крупных таксономических групп (Биттерман, 1973; см. 3.3.1). При последовательных переделках сигнального значения стимула формирование каждого следующего УР у многих млекопитающих (в том числе у крыс) и некоторых видов птиц ускоряется, т.е. число ошибок при каждом новом сигнальном значении стимулов постепенно и достаточно заметно снижается. В отличие от них у рыб такого ускорения практически не происходит. Таким образом, способность уловить общее правило, лежащее в основе теста, проявляется у некоторых животных с относительно примитивным мозгом (крысы, голуби).
Формирование установки на обучение (см. 3.3.3) также позволило выявить различия в способности к обучению у животных разных видов, коррелирующие с уровнем развития мозга (рис. 1).
Как видно из рисунка, у большинства видов приматов установка на обучение формируется после выработки 150—200 дифференциро-вок. Иными словами, в этот период доля правильных выборов уже при втором предъявлении новых стимулов (т.е. без дополнительного обучения) доходит до 90%. Несколько больше таких серий обучения требуется беличьим саймири, еще больше — мармозеткам и кошкам. В отличие от них у крыс, кур, голубей и белок правильные ответы при втором предъявлении новой пары стимулов превышали случайный уровень не более чем на 10—15% даже после выработки 1500 различных дифференцировок. Для крыс этот тест оказался менее доступным, чем многократная переделка УР.
Врановые птицы — американские сойки (Суапосilla cristata) и вороны (Corvus brachyrhynchos), а также скворцы-майны (Gracula religiosa) по скорости формирования установки на обучение превосходят кошек и саймири и приближаются к мартышковым обезьянам. Для них характерны также сходные стратегии решения теста (см. ниже).
Скорость формирования установки на обучение соответствует уровню организации мозга: низкая — у грызунов, выше — у хищных млекопитающих и очень высокая — у приматов в целом.
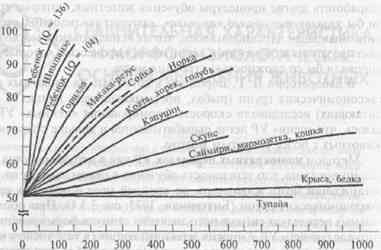
Рис. 1. Сравнительная характеристика способности животных разных видов к формированию установки на обучение.
По оси ординат — доля (%) правильных выборов при второй пробе, по оси абцисс — предъявление задачи. По Hodos, Campbell, 1979, с дополнениями.
Вместе с тем внутри отряда приматов существуют определенные различия по этому показателю. Наиболее успешно формируют «установку» человекообразные обезьяны, причем шимпанзе опережают в этом отношении других антропоидов, включая горилл, и даже детей с низким коэффициентом интеллектуального развития. Это служит одной из важных иллюстраций широко распространенного представления об исключительной близости шимпанзе к человеку (Rumbaugh et al., 2000).
Наряду со столь очевидными различиями показателей решения, которые обнаружены у шимпанзе и грызунов, во многих случаях животные с заведомо по-разному организованным мозгом (например, кошки и макаки) демонстрируют сходные количественные показатели формирования установки. Однако различия между ними выявляются четко, если обратиться к «качественному» анализу, т.е. к сопоставлению стратегий в решении этого теста. Такой анализ провел Д. Уоррен (Warren, 1977). Кошки действовали достаточно примитивно. Если при первом предъявлении новой пары стимулов кошка случайно выбрала «правильный» стимул, то дальше действовала безошибочно, т.е. придерживалась стратегии «преуспел — делай так же» (win-stay). При неудачном выборе кошка не могла адекватно использовать полученную информацию и в следующий раз выбирала наугад, не используя стратегию «проиграл — измени тактику» (lose-shift), а дальше обучалась по принципу «проб и ошибок».
В таких же опытах макаки-резусы вели себя иначе. Даже если при первом предъявлении новой пары стимулов их выбор был неудачным, то при следующих пробах они практически всегда действовали правильно. Иными словами, «неуспех» воспринимался макаками не как ошибка, после которой надо снова пробовать, а как основание для смены стратегии выбора, т.е. они далее действовали по правилу: «проиграл — измени тактику». К тому же, в отличие от кошек, макаки могли гибко переходить от одного правила к другому. Это означало, по-видимому, что они способны полностью уловить принцип задачи. Такую же более совершенную стратегию формирования «установки» проявили представители врановых птиц — сойки: они реагировали на новые стимулы правильно независимо от того, был ли выбор в первой пробе положительным или отрицательным (Kamil, 1988).