Функциональная архитектура зрительной коры
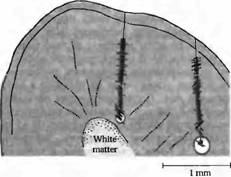
Рис. 3. Оси ориентации рецептивных полей нейронов по мере погружения электрода вглубь коры мозга кошки. Все последующие клетки демонстрируют одинаковую ориентацию оси, что показано в виде угла, под которым располагается штриховая полоска по отношению к линии проникновения электрода. Погружение электрода справа идет с большим отклонением от перпендикуляра, следовательно, при пересечении им нескольких колонок часто меняется и ориентация оси. Расположение каждой клетки определяется при помощи нанесения небольших повреждений по мере погружения электрода и при завершении погружения (показано кружком). На основании этих данных производится реконструкция хода электрода. Подобные эксперименты позволили установить, что клетки со сходными свойствами у кошек и обезьян организованы в виде колонок, идущих под определенными углами к поверхности коры.
Ориентационные колонки
Какие другие функциональные объединения возможны для зрительных нейронов в зоне V1?
Ориентационные предпочтения простых и комплексных клеток. Таким образом, логичен вопрос: является ли данная особенность систематически повторяющейся в зрительной коре? Репрезентативный эксперимент, отвечающий на этот вопрос, представлен на рис. 3. Микроэлектрод погружается перпендикулярно поверхности вглубь серого вещества в зоне V1 мозга кошки. Каждый штрих на этом рисунке показывает расположение одной клетки и предпочтительную ориентацию ее рецептивного поля при предъявлении визуальных раздражителей в виде полосок. После погружения электрода в контрольной точке клетки локально повреждаются посредством пропускания электрического тока. Расположение каждой регистрируемой клетки затем реконструируется на основании этих контрольных точек (конечная точка показана кружком в конце пути электрода). Как показано с левой стороны рисунка, первые (начиная с поверхности) 38 клеток наиболее эффективно активировались полосками, расположенными под углом около 90 градусов к вертикальной оси. После погружения на глубину около 0,6 мм ось ориентации рецептивного поля клеток сместилась примерно на 45 градусов. При другом погружении, показанном справа, с более острым углом к поверхности коры, каждая последующая клетка имеет разные варианты ориентации оси. При таком наклонном проникновении оси полей меняются по мере прохождения через разные колонки, имеющие отличающуюся ориентацию. Ориентационные колонки получают сигналы от клеток, рецептивные поля которых значительно перекрываются на поверхности сетчатки.
Информация об организации ориентационных колонок в зрительной коре обезьян и кошек была впервые получена при проведении именно косых (тангенциальных), а не перпендикулярных погружений электрода в глубь коры. При продвижении электрода на каждые 50 мкм в горизонтальном направлении вдоль коры ориентация оси рецептивного поля меняется примерно на 10 градусов, иногда такие последовательные смены ориентации достигают в конечном итоге 180 градусов. Колонки ориентации (20-50 мкм) гораздо уже, чем глазодоминантные колонки (250-500 мкм). Первая анатомическая демонстрация ориентационных колонок, основанная на их разной функциональной активности, была произведена Соколовым, который использовал 2-деоксиглюкозу, хорошо захватывающуюся активными клетками. В результате метаболически активные клетки становились радиоактивными и их расположение можно было видеть на ауторадиограммах. У обезьян и кошек, которым показывали горизонтальные или вертикальные полосы, «полоски» радиоактивности совпадали с расположением колонок вертикальной или горизонтальной ориентации, различимых в коре без всякой окраски. Организация ориентационных колонок изучалась также при помощи оптических методов на живых животных. Примером может служить эксперимент Бонхоеффера и Гринвальда. Представление зрительных стимулов различной ориентации приводило к активности в различных кортикальных зонах. Ответы на разные ориентации показаны разными цветами. Удивительной является организация ориентационных колонок по отношению друг к другу. Сначала их организация кажется достаточно беспорядочной. Однако при тщательном исследовании можно заметить определенные центры в виде пестрого «волчка», в котором сходятся отдельные точки всех ориентаций. Отсюда клетки, ответственные за определенную ориентацию, расходятся в удивительно правильном порядке. Некоторые волчки ориентированы по часовой стрелке, некоторые — против. Таким образом, ориентация в коре представлена скорее в радиальном, чем в линейном виде. Каждый угол ориентации уникален (возникает только один раз) в пределах цикла, вертикальные и горизонтальные контуры занимают несколько большую площадь коры. На каждом квадратном миллиметре коры находятся один или два таких центра, равноудаленных друг от друга. Интересно то, что подобный способ организации коры был предложен ранее, на основании теоретических предпосылок.
Связь между колонками глазного доминирования и ориентационными колонками
При помощи методов оптической микроскопии была также выявлена взаимосвязь между ориентационными колонками и колонками глазного доминирования. Структура корковой активности регистрировалась сначала при стимуляции одного глаза, а затем определялась снова при помощи серии ориентированных световых полос. Результаты одного такого эксперимента .Каждый контур, имеющий свою ориентацию, показан в виде отдельной цветной линии (и изоориентированного контура), зоны глазного доминирования показаны в виде светлых или темных участков. Волчки ориентации четко видны в виде сходящихся изоориентированных контуров, при этом несколько контурных линий между различными волчками обычно пересекают границу глазного доминирования. Таким образом, получается, что большинство ориентационных зон оказывается расщеплено на контралатеральную и ипсилатеральную половинки, получая информацию от обоих глаз с данного места зрительного поля. На самом деле, центр каждого волчка стремится оказаться наиболее близко к центру глазного доминирования, а контуры изоориентации имеют свойство пересекать границы глазного доминирования под определенными углами .
Параллельная обработка информации о форме, движении и цвете
Колонки зрительного доминирования и ориентационные колонки представляют собой только два типа возможной функциональной организации нейронов зрительной коры. Направление движения, пространственная частота (в основном зависящая от размеров рецептивного поля), несоразмерность изображения (важная детерминанта для восприятия глубины изображения) также обеспечиваются функциональными образованиями зрительной коры, имеющими вид колонок. Возникает логичный вопрос: каким образом выполняются все необходимые виды анализа изображения для каждой точки в зоне коры, куда проецируется ретинотопическая карта? Эффект «смешивания» функциональных колонок, дает частичный ответ на этот вопрос. В самом деле, задолго до использования оптических методов для поиска подобных взаимосвязей, Хьюбель и Визель предложили концептуальную схему того, что они назвали «сверхколонка» (hypercolumn). Под этим термином они понимали такое объединение нейронов, в котором представлены все возможные ориентации соответствующих областей зрительных полей обоих глаз. Согласно этому представлению соседние сверхколонки анализируют информацию таким же образом, как и соседние перекрывающиеся части зрительных полей. Тем не менее остается пока трудной задачей объединить эту точку зрения с данными современных методов анализа изображения.