Функциональная архитектура зрительной коры
Проекции в зрительную зону 2 (V2)
При окрашивании на цитохромоксидазу зоны V2 выявляется паттерн, отличный от того, который виден в зоне V. Окрашивание происходит в виде серии тонких и толстых полосок, перемежающихся с более бледными зонами с меньшей активностью фермента (рис. 4. Эти параллельные полоски идут под определенными углами от границы между зонами V1 и V2. После введения фермента пероксидазы хрена в окрашенные зоны V1, он захватывается терминалами аксона и транспортируется ретроградно, показывая, что нейрон, отростки которого проецируются в пределах зоны «пятна», расположен в пределах тонкой полосы окрашивания зоны V2 зрительной коры. Эти связи взаимны: при инъекции в тонкие полоски возникает окрашивание в области «пятен». И наоборот, области между «пятнами» образуют связи с бледными широкими полосами, которые получают в основном информацию от больших клеток из слоев 4В и 4С. Примечательно, что это функциональное подразделение может быть выявлено даже на молекулярном уровне: моноклоналъные антитела Cat-301 в основном маркируют крупноклеточные пути в пределах зрительной коры обезьяны.

Рис. 4. «Пятна» в зоне V1 и «полоски» в зоне V2 зрительной коры обезьян, окрашенной на цитохромоксидазу. Пятна организованы в виде узора «в горошек». Можно различить четкую границу между зонами V1 и V2 . По этой линии «пятна» переходят в «полоски», толстые и тонкие, идущие под определенным углом к границе.
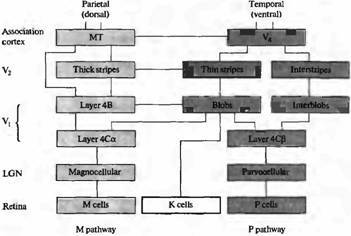
Рис. 5. Схематическая организация М-, Р- и К-кана-лов, направляющихся в зрительную кору. Функционально различные слои ЛКТ направляют отростки к различным слоям в V1. К-слои проецируются в «пятна» в слое 2 и 3. М- и Р-слои области 4С могут взаимодействовать предпочтительно с областями «пятна» и с областями, расположенными между пятен в слоях 2 и 3. «Пятна» проецируются преимущественно в тонкие полоски в V2 . Тонкие полоски проецируются в V4. Толстые полосы в V2 получают информацию от слоя 4В в V и проецируются в ассоциативную зону МТ (V5). M-каналы проецируются в дорзальную (теменную) зрительную кору, где происходит анализ движения. Р-каналы проецируются предпочтительно в зону V4, где осуществляется обработка информации о цвете.
Ассоциативные зоны зрительной коры
Значительные усилия были потрачены для понимания процесса переработки информации в М- и Р-каналах зрительной коры. Подобные исследования были вызваны функциональными и анатомическими различиями ганглиозных клеток сетчатки и нейронов коленчатого тела (рис. 4). Дополнительная информация обеспечивалась психофизическими экспериментами, исследованием строения головного мозга, изучением пациентов с различными мозговыми повреждениями. Все это привело к предположению, что различные типы анализа зрительной информации производятся в физически изолированных регионах мозга, в так называемых ассоциативных зонах коры. Дорзальный или париетальный тракт (называемый так потому, что соответствующие зоны коры находятся в теменной доле, расположенной более дорзально по отношению к первичной зрительной коре), как считают, является важным звеном в оценке движения и пространственного взаиморасположения форм — свойств, сходных со свойствами М-каналов, которые мы уже описали. Повреждения дорзального, париетального тракта выражаются в игнорировании участка зрительного поля и нарушении видеомоторной ориентации. Повреждения в области височной коры (вентральнее V1) уменьшают способность к зрительной идентификации объектов, их цветов и тонких деталей — отражая свойства Р-каналов.
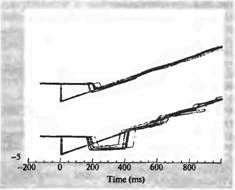
Рис. 6. Организация зрительного движения в зоне МТ Обезьяна была обучена отслеживать взглядом движущиеся предметы (движение стимула показано светлой серой линией). На верхней записи показано положение глаз обезьяны относительно движущегося объекта. После начального резкого быстрого движения предмет проецируется на зрительной ямке (быстрое отклонение глаза вниз), затем глаз точно следует за движением предмета. Однако после введение нейротоксина в область МТ (нижняя запись) первое движение является избыточным и «проскакивает» цель. Дальнейшее слежение идет медленнее, чем это необходимо, как будто была неправильна рассчитана скорость движения предмета.
Детекция движения и зона МТ
Если попросить дать краткое определение, что такое зрение, большинство из нас скажет о восприятии объектов, включающем анализ тонких деталей, примерно как это происходит во время чтения этих слов. Однако не менее важной функцией зрения является анализ движения, хотя эта функция и остается в основном непроизвольной и подсознательной (за исключением тех, кто из-за окуломоторного расстройства не может автоматически управлять перемещением зрительного поля). Как уже было описано, движение анализируется крупноклеточно-париетальной частью зрительного тракта (рис. 4). Нейроны крупноклеточного пути чувствительны к движущимся стимулам, это их свойство сохраняется в зонах V1 и V2 (в пределах толстых полос), через которые этот путь проходит, направляясь к средневисочной ассоциативной коре (зона МТ, или V5). Зона МТ также имеет ретинотопическое строение. Нейроны в этой области чувствительны к скорости и направлению движения стимула, а также объединены в группы в виде колонок со сходными предпочтительными направлениями движения. Когда небольшие области МТ химически повреждаются при помощи нейротоксина, у обезьяны нарушается способность воспринимать характер движения точек в соответствующих областях зрительного поля, хотя чувствительность к контрастности изображения при этом не меняется.
Зона МТ и зрительное слежение
Зона МТ играет важную роль в процессе зрительного слежения. Это было показано рядом экспериментов, в которых обезьяну обучали следить глазами за движущимся предметом (рис. 5). Нормальный паттерн движения глаз показан на верхней записи на рис. 5. При этом при движении объекта (траектория начинается от точки 0) глаза фокусировались на нем при помощи быстрого, отрывистого движения (с отклонением вниз через 200 миллисекунд), а затем при помощи аккуратного слежения предмет удерживался в области зрительной ямки. После введения небольшого количества нейротоксина (иботеновой кислоты) в зону МТ, в область представительства центральной ямки, способность обезьяны следить за движущимся объектом значительно нарушалась. В частности, после первого быстрого движения дальнейшая скорость слежения была значительно ниже скорости перемещения объекта. Нарушено было также и само начальное установочное движение глаз (рис.5, нижний пробег). Таким образом, повреждение зоны МТ приводило к тому, что животное теряло способность адекватно отслеживать движение объекта.
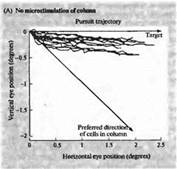
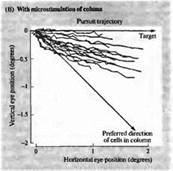
Рис. 6. Изменение направления движения глаза при электрической стимуляции в области МТ (А) Запись движения глаза в ответ на движущийся зрительный стимул. До этого микроэлектрод был введен в область зоны МТ, и было определено предпочтительное направление движения для клеток этой области, которое отличается от направления движения данного стимула (В). При электрической стимуляции зоны МТ результирующий вектор движения отклоняется в сторону. Эти данные дают основания предполагать, что зрительные движения вычисляются как суммарный вектор нескольких предпочтительных направлений в зоне МТ.