Что общего в иммунитете растений и животных
Иммунные молекулы
Строение антител - иммуноглобулинов - хорошо известно: две полипептидные цепи - тяжелая (H, молекулярная масса ~50 кДa) и легкая (L, молекулярная масса ~25 кДa), соединенные дисульфидными связями. Каждая цепь содержит чередующиеся константные (С) и вариабельные (V) участки. Огромное разнообразие антител в организме, позволяющее узнавать множество чужеродных антигенов, обусловлено уникальным генетическим контролем их синтеза.
Продукты генов устойчивости растений названы R-белками (от англ. resistance - устойчивость). Структуру первых трех R-белков (из томатов, арабидопсиса и табака) установили сравнительно недавно - в 1993 г. С тех пор их описано несколько десятков.
В состав R-белков входит несколько структур, обеспечивающих, с одной стороны, взаимодействие с лигандом (элиситором) и с молекулами-мишенями (в частности, с ДНК), а с другой - передачу сигнала на другие молекулы - циклические нуклеотиды, протеинкиназы и др. Полипептидные цепи R-белков образованы комбинациями фрагментов, что и обеспечивает растениям из разных классов и порядков устойчивость к вирусам, бактериям, грибам и даже нематодам. Следовательно, эти структуры возникли очень давно, до расхождения предковых растений на современные классы и порядки, выполняли иные, более общие, функции, связанные с рецепцией и передачей сигнала от возбужденного рецептора к генам. Значит, их иммунная функция вторична.
Гены устойчивости растений, контролирующие синтез R-белков, расположены на хромосомах неслучайно. Широко распространены три типа локализации: одиночное диаллельное (один локус с двумя аллелями, контролирующими восприимчивость и устойчивость, которая, как правило, доминантна); одиночное мультиаллельное (один локус с множеством кодоминантных аллелей, контролирующих устойчивость к разным видам и расам патогена); сцепленное (большое число ди- или мультиаллельных локусов сцеплены и образуют блок фенотипически сходных генов, определяющих устойчивость к одной или нескольким болезням). Вот некоторые примеры.
Ген устойчивости риса к бактерии Xanthomonas oryzae Xa21 входит в мультигенное семейство, состоящее по крайней мере из восьми членов, которые картируются в одном локусе на хромосоме 11. На коротком плече V-й хромосомы ячменя находится блок из пяти локусов, определяющих устойчивость к мучнистой росе; в одном из локусов - Mla - картировано около 20 кодоминантных аллелей. Устойчивость кукурузы к расам возбудителя северной ржавчины Puccinia sorgi контролируют пять локусов. Два из них - Rpl и Rp5 - тесно сцеплены, причем в локусе Rpl картировано 14 аллельных и тесно сцепленных генов. Гены устойчивости томатов к кладоспориозу (возбудитель - гриб Cladosporium fulvum) образуют несколько групп сцепления. Тесно сцеплены гены Cfl, Cf-4 и Сp, расположенные на коротком плече хромосомы 1, а также Cf-2 и С-5 на коротком плече хромосомы 6. Последние, вероятно, аллельны, так как не рекомбинируют у гибридного потомства.
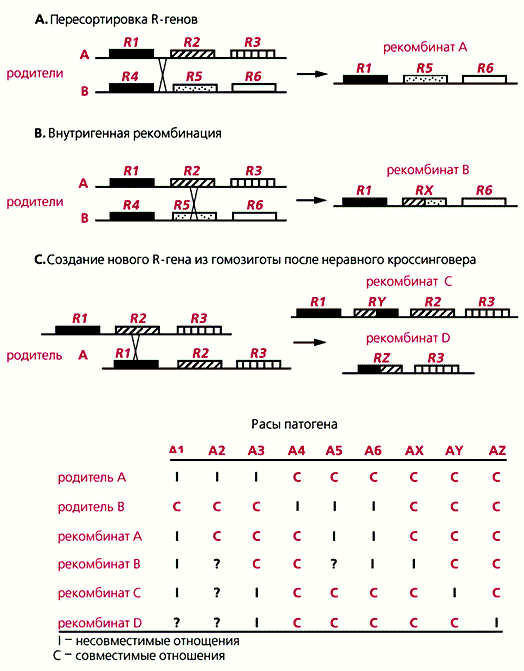
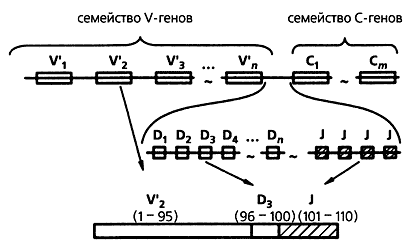
Схема участка хромосомы, включающего семейства вариабельных, V, и константных, C, генов (вверху). Численность V-генов в отдельных семействах варьирует от 50 до 1000, а C-генов - от 1 до 9. Внизу - сборка V-гена: V-фрагмент - контролирующий первые 95 аминокислот, D - следующие за ним от 2 до 10, J - концевой фрагмент V-гена, контролирующий от 5 до 10 концевых аминокислот. Районы ДНК, разделяющие V-D и D-J фрагменты, вырезаются и утрачиваются при сборке гена.
Возникновение кластеров сцепленных генов, функционально и, вероятно, структурно сходных, обусловлено внутри- или межгенными обменами участков ДНК, имеющих прямые (аbсd - а’b’с’d’) или инвертированные (аbсd - d’с’b’a’) повторяющиеся последовательности. Нити ДНК двух родителей в мейозе могут спариваться по гомологичным последовательностям, которые находятся в негомологичных участках хромосом. Это приводит к неравному кроссинговеру. В результате у одного родителя участок хромосомы теряется (делетируется), а у второго - удваивается (дуплицируется), т.е. вместо одного гена появляется два сцепленных.
С этой точки зрения наибольший интерес представляет LRR-область R-белков с многочисленными повторяющимися последовательностями. Благодаря точковым мутациям, делециям, инверсиям такая структура обеспечивает генетическую реорганизацию. Экспериментально показано, что мутации, изменяющие реакцию на заражение авирулентными расами патогенов, картированы в LRR-области.
Большую роль в увеличении генетического разнообразия семейств генов устойчивости могут играть разрывы кодирующей части ДНК, вызванные внедрением мобильных генетических элементов. Так, у риса в локусе белка Ха21 обнаружены транспозоны Truncator и Retrofit, приводящие к разрывам и образованию самостоятельных рамок считывания. Этот белок сочетает свойства двух белков томата - мембранного рецептора и фермента протеинкиназы. Возможно, возникновение генов С/9 и Pto обусловлено разрывом предшественника, подобного гену Ха21.
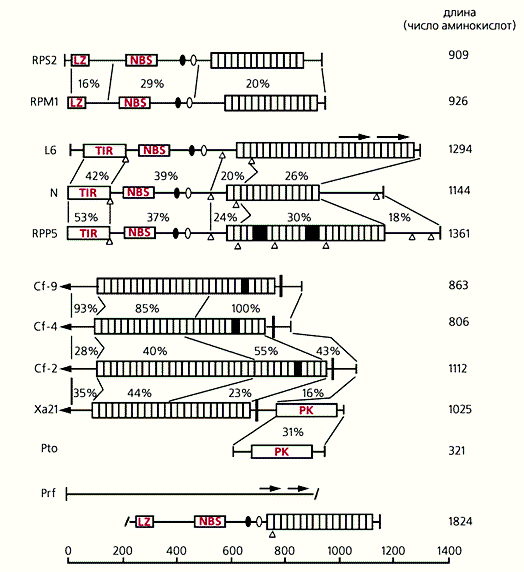
Схема строения R-белков. Заштрихованы С-концевые домены, которые содержат повторяющиеся последовательности, богатые лейцином (LRR-области). Каждый повтор состоит из 23-24 аминокислот. Эта структура, встречающаяся во многих белках эвкариот, осуществляет белковые взаимодействия, т.е. служит рецептором, связывающимся с лигандом - элиситором патогена. NBS (Nucleotide Binding Sites) - сигнальная область, связывающаяся с АТФ и ГТФ, вследствие чего она может активировать киназы или сигнальные G-белки. LZ (Leucine Zipper Region) - область лейциновой “застежки”, которая участвует в формировании спирализованных структур, ответственных за димеризацию или специфическое взаимодействие с другими белками. TIR (Toll/Interleukin-1 Resistance) - область гомологии с цитоплазматическим доменом Toll-белка дрозофилы и рецептором интерлейкина-1 млекопитающих. Возможно, TIR-область растительных R-белков несет сходные функции. РК - серин-треониновая протеинкиназа - фактор активации транскрипции и других сигнальных путей. Слева обозначены гены растений, кодирующие R-белки: RPS и RPM - гены устойчивости арабидопсиса к бактериям из рода Pseudomonas; L - ген устойчивости льна к ржавчине; N - ген устойчивости табака к вирусу табачной мозаики; RPP - ген устойчивости арабидопсиса к ложной мучнистой росе; Cf - гены устойчивости томатов к грибу Cladosporium fulvum; Xa - ген устойчивости риса к бактерии Xanthomonas oryzae; Pto и Prf - система генов, контролирующих устойчивость томатов к бактерии Psiulomonas tomato. Справа - содержание аминокислот в белках. Внутренние цифры - проценты идентичных последовательностей в специфических областях. Стрелки указывают направления транскрипции, а треугольники - положение интронов.
Разные типы рекомбинаций (A, B, C) на хромосомном сегменте, содержащем локусы устойчивости растений к болезням (А – С). Внизу - гипотетические изменения устойчивости к расам патогенного микроорганизма, возникающие в результате рекомбинаций.