Витамины В12 и В15
Глутаматмутаза
Фермент, катализирующий превращение L-глутамата в L-трео-β-метиласпартат, был выделен, из Clostridium tetanomorphum. Показано, что многочисленные фотосинтезирующие микроорганизмы также содержат глутаматмутазу. Фермент, кроме аденозилкобаламина, нуждается в SH-соединении, однако, в отличие от диолдегидратаз и других аналогичных ферментов, для проявления каталитической активности не требуются одновалентные катионы. Фермент высокоспецифичен в отношении структуры субстратов. Ни аналоги глутаминовой кислоты, ни аналоги β-метиласпарагиновой кислоты (как, например, β-этиласпартат) не являются субстратами мутазы.
 Опыты по очистке фермента позволили установить субъединичную структуру и этого аденозилкобаламин-зависимого фермента. Были получены в гомогенном состоянии две субъединицы, которые были названы S- и Е-белками. Каждый из белков не обладал порознь активностью. Рекомбинация их и взаимодействие с коферментом приводили к восстановлению активности. Первым был очищен Е-белок. Определение молекулярного веса показало, что это довольно большой белок с мол. массой около 128 000. В отличие от ферментов, рассмотренных выше, добавление Кобаламина не защищало Е-компонент глутаматмутазы от инактивации в растворе. Е-компонент связывал 1 моль аденозилкобаламина, а в присутствии семикратного избытка S-компонента дополнительно связывал еще один моль кофермента.
Опыты по очистке фермента позволили установить субъединичную структуру и этого аденозилкобаламин-зависимого фермента. Были получены в гомогенном состоянии две субъединицы, которые были названы S- и Е-белками. Каждый из белков не обладал порознь активностью. Рекомбинация их и взаимодействие с коферментом приводили к восстановлению активности. Первым был очищен Е-белок. Определение молекулярного веса показало, что это довольно большой белок с мол. массой около 128 000. В отличие от ферментов, рассмотренных выше, добавление Кобаламина не защищало Е-компонент глутаматмутазы от инактивации в растворе. Е-компонент связывал 1 моль аденозилкобаламина, а в присутствии семикратного избытка S-компонента дополнительно связывал еще один моль кофермента.
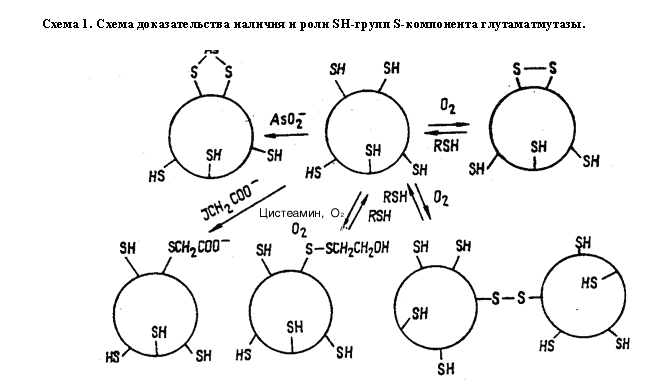
Компонент S после очистки, как оказалось, обладал намного меньшей мол. массой 17000 и, по-видимому, содержал важные для проявления ферментативной активности SH- группы. Титрование S-белка сульфгидрильными реагентами показало, что на 1 моль белка приходится пять SH-групп. Примечательной особенностью компонента была его способность к димеризации в присутствии О2. Расщепление димера осуществлялось обработкой последнего каким-либо RSH-соединением. Это свидетельствовало об образовании межмолекулярного дисульфидного мостика. Инактивация S-белка с помощью AsO2- доказывала наличие в активном центре по крайней мере одной из двух вицинальных тиольных групп. Превращение SH-групп S-белка представлены на схеме 3.
Метилмалонил-СоА-мутаза
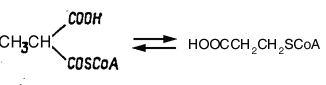
Другой аденозилкобаламин-зависимый фермент, осуществляющий перегруппировку углеродного скелета метилмалонил-СоА в сукцинил-СоА был также сначала выделен из микроорганизмов, а затем из тканей млекопитающих. Оказалось, что этот кобаламин-зависимый фермент выполняет метаболически важную роль на пути превращения пропионил-СоА в сукцинил-СоА. Схема этого участка метаболизма включает 3 фермента: биотин-зависимую карбоксилазу, рацемазу, превращающую D-метилмалонил-СоА в L-изомер и рассматриваемую кобаламин-мутазу. После очистки метилмалонил-СоА-мутазы оказалось, что это субъединичный фермент с мол. массой 124000, расщепляющийся на два компонента с мол. массой 61 000 и 63000. Выделенная из печени овцы метилмалонил-СоА-мутаза представляет собой окрашеный в оранжевый цвет белок с мол. массой 165000. Фермент связывал 1 моль аденозилкобаламин на 75000.
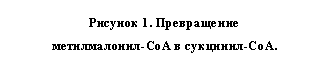
2-метиленглутаратмутаза

 Следующий сходный по действию Аденозилкобаламин-зависимый фермент-это 2-метиленглутаратмутаза, катализирующая обратимое превращение между 2-метиленглутаратом и 2-метилен-З-метилсукцинатом. Фермент был выделен из микробиологических источников при выращивании Clostridium на средах, содержащих никотиновую кислоту. Определение молекулярной массы частично очищенного препарата дало величину 170000. Обработка йодацетатом приводит к потере активности, что, очевидно, свидетельствует о наличии важных для катализа SH-групп. Действие AsО2-, однако, не выявило присутствия вицинальных дитиольных групп.
Следующий сходный по действию Аденозилкобаламин-зависимый фермент-это 2-метиленглутаратмутаза, катализирующая обратимое превращение между 2-метиленглутаратом и 2-метилен-З-метилсукцинатом. Фермент был выделен из микробиологических источников при выращивании Clostridium на средах, содержащих никотиновую кислоту. Определение молекулярной массы частично очищенного препарата дало величину 170000. Обработка йодацетатом приводит к потере активности, что, очевидно, свидетельствует о наличии важных для катализа SH-групп. Действие AsО2-, однако, не выявило присутствия вицинальных дитиольных групп.
Ферменты, трансформирующие α, ω-диаминокислоты
Еще одна группа аденозилкобаламин-зависимых ферментов была выявлена при исследовании микроорганизмов, растущих на L-лизине. Эти ферменты катализировали перенос NH2-гpyппы от концевого углеродного атома в диаминокислотах к соседнему атому углерода. Недавно описан аденозилкобаламин-зависимый фермент, который осуществляет перенос NH2-группы из α- в β-положение аминокислоты лейцина.
Рассмотрим более подробно аденозилкобаламин-зависимые ферменты, трансформирующие α, ω-диаминокислоты. Эти ферменты выделены из различных штаммов Clostridium.
Важно, что все три фермента в высшей степени специфичны для каждого из субстратов. Замена субстрата для данного фермента на субстраты других аминомутаз не позволяет осуществлять перенос аминогруппы.
D-α-Лизинмутаза выделена из С. sticklandi, мол. масса 250000. В процессе очистки фермента не происходило отщепления аденозилкобаламина, что свидетельствует о большой прочности связи комплекса апофермент-кофермент. Аналогично от L-β-лизинмутазы в процессе очистки аденозилкобаламин также не отделялся. Это тетрамерный фермент с мол. массой 160000; его субъединичная структура была подтверждена после того, как удалось выделить компоненты с мол. массой 32000 и 52000. Оказалось, что для обеспечения нормальной каталитической активности обеих мутаз необходим белок с мол. массой 60000. Как видно, аминомутазы сохраняют способность защищать в полностью реконструированном виде важные для биокатализа SH-группы. Это свойство характерно для большинства Аденозилкобаламин-ферментов.
Орнитинмутаза также представляет собой фермент с прочно связанным аденозилкобаламином, который не отщепляется при очистке. Очищенный фермент имел мол. массу 170 000 и диссоциировал на две субъединицы с мол. массой 90000. Этот фермент не требовал для своей активации какого-либо SH-белка, однако сам содержал важные для биокатализа SH-группы, вследствие чего ингибировался под действием тиоловых ингибиторов.
Заслуживает особого внимания прочность, с которой связан аденозилкобаламин во всех трех ферментах. При обработке их внутренним фактором ферменты инактивировались. Сравнительное исследование трех аминомутаз показало, что в дополнение к аденозилкобаламину для проявления их активности требуются: для D-α- и L-β-мутаз ионы Mg2+ и одновалентные катионы, причем для D-α-лизинмутазы-это К+, Rb+ и NH4+ (фермент ингибируется при добавлении Na+ и Li+). L-β-лизин-мутаза в разной степени активна в присутствии Li+, Na+, К+, Rb+, Cs+ и. NH4+. Для проявления активности орнитинмутазы добавления ионов металлов вообще не требуется.
Поразительное отличие аминомутаз от всех остальных кобаламин-зависимых ферментов состоит в том, что вторым обязательным коферментом для них является пиридоксальфосфат, который обеспечивает перенос ω-NH4-гpyппы в форме пиридоксамина или пиридоксальдимина. Очищенная орнитинмутаза неактивна до тех пор, пока к реакционной смеси не будет добавлен пиридоксальфосфат (PLP). Очевидно, что PLP входит в активный центр фермента и, по-видимому, связан в виде основания. Шиффа с ε-NН4-группой остатка лизина белка.