Витамины В12 и В15
Участие витамина B12 в биохимических
восстановительных процессах
Утверждали, что витамин В12 помимо действия на сульфгидрильные соединения поддерживает в восстановленном состоянии другие важные вещества. Так, Уилл и сотр. установили, что в плазме больных пернициозной анемией содержание аскорбиновой кислоты понижено; кроме того, при инъекции таким больным аскорбиновой кислоты она быстро окисляется в дегидроаскорбиновую. После лечения витамином B12 эти явления исчезают, а инъекции аскорбиновой кислоты ведут к повышению ее концентрации в плазме. Чоу и сотр. нашли, что в печени крыс с недостаточностью витамина В12 общее содержание дифосфопиридиннуклеотида повышено, но количество его восстановленной формы (ДПН-Н) понижено. Ненормально высокое отношение ДПН/ДПН-Н снижалось вдвое после введения витамина B12.
Было высказано предположение, что витамин B12 способен играть роль восстановителя, когда его трехвалентный кобальт восстановлен до двухвалентного состояния. Однако нужны сильные восстановители, чтобы вызвать эту реакцию, которая в присутствии атмосферного кислорода идет в обратном направлении. Предположение о том, что соединение с белком могло бы сдвинуть окислительно-восстановительный потенциал в область физиологических величин, не вполне убедительно, так как способность связывать белок после восстановления, возможно, утрачивается.
Биосинтез метионина и серина
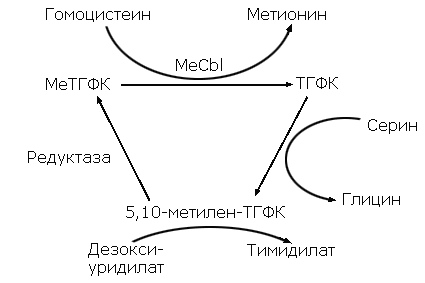
Метилкобаламин участвует в реакциях синтеза метионина в качестве кофактора. Заключительным этапом синтеза метионина у бактерий, грибов, высших растений и животных состоит в переносе метильной группы от СН3-ТГФК к сульфгидрильной группе гомоцистеина.
Существует два типа ферментов, осуществляющих синтез метионина – первый, независимый от кобаламинов, может использовать в качестве донора метильной группы только триглутаматную форму СН3-ТГФК. Второй тип ферментов, зависимый от кобаламиновых кофакторов, может использовать как моно-, так и триглутаматную форму СН3-ТГФК. Для активации ферментов второго типа, кроме того, требуется S-аденозилметионин (S-AdoMet). Последний необходим для первоначального метилирования кобаламина.
Мутант Е. coli, используемый для определения витамина В12, способен так же хорошо расти и при добавлении к минимальной питательной среде метионина, только для оптимального роста требуется примерно в 10000 раз больше метионина, чем витамина. Очевидный вывод, что в клетках этого организма витамин действует как катализатор синтеза метионина, был подтвержден экспериментально. Однако для любого другого микроорганизма, нуждающегося в витамине B12, этот витамин не может быть заменен метионином, так что он, очевидно, осуществляет у этих организмов какую-то дополнительную функцию.
|
Скармливаемый предшественник |
Радиоактивность метильного углерода метионина, μ с/г-атом |
Повышение биосинтеза метионина, % | |
|
В присутствии вит. В12 |
В отсутствие вит. В12 | ||
|
При неограниченном потреблении пищи | |||
|
α-14С-Глицин (2%) β-14С-Серин (0,7%) 14С-Формиат натрия (0,1%) |
156 81,5 33,8 |
95,5 47,4 20,0 |
63 72 69 |
|
При ограниченном потреблении пищи | |||
|
14С-Формиат натрия (0,1%) |
32,0 |
28,6 |
12 |
Таблица 1. Влияние витамина В12 на биосинтез метионина.

 Ранние эксперименты с изучением роста цыплят и крыс также показали, что витамин B12 снижает потребность в метионине, особенно при введении гомоцистеина. Сначала это было истолковано как действие витамина на трансметилирование, т. е. на передачу лабильной метильной группы от холина или бетаина к гомоцистеину с образованием метионина. Точно так же витамин B12 может, по крайней мере частично, заменять холин для цыплят, крыс и поросят-сосунков. Ряд исследований (некоторые из них с использованием 14С) показал, что витамин B12 не оказывает никакого влияния на трансметилирование, но участвует в прямом синтезе лабильной метильной группы из более окисленных предшественников – таких, как формиат, α-углерод глицина или β-углерод серина. Трудности истолкования результатов, получаемых на интактных животных, хорошо иллюстрирует таблица 1.
Ранние эксперименты с изучением роста цыплят и крыс также показали, что витамин B12 снижает потребность в метионине, особенно при введении гомоцистеина. Сначала это было истолковано как действие витамина на трансметилирование, т. е. на передачу лабильной метильной группы от холина или бетаина к гомоцистеину с образованием метионина. Точно так же витамин B12 может, по крайней мере частично, заменять холин для цыплят, крыс и поросят-сосунков. Ряд исследований (некоторые из них с использованием 14С) показал, что витамин B12 не оказывает никакого влияния на трансметилирование, но участвует в прямом синтезе лабильной метильной группы из более окисленных предшественников – таких, как формиат, α-углерод глицина или β-углерод серина. Трудности истолкования результатов, получаемых на интактных животных, хорошо иллюстрирует таблица 1.
 При авитаминозе В12 сильно ухудшается аппетит и наблюдаемые результаты часто могут быть обусловлены просто пониженным потреблением пищи по сравнению с контрольными животными. Эту неясность можно устранить, ограничив потребление пищи контрольными животными до уровня,
При авитаминозе В12 сильно ухудшается аппетит и наблюдаемые результаты часто могут быть обусловлены просто пониженным потреблением пищи по сравнению с контрольными животными. Эту неясность можно устранить, ограничив потребление пищи контрольными животными до уровня,  характерного для авитаминозных животных (метод "парного кормления"). Подтверждение данных для поросят и цыплят в опытах с 14С-формиатом и 14С-серином получили Джонсон и сотр. Однако при использовании меченого формальдегида результат оказался неожиданным: интенсивность включения метки в метильные группы метионина и холина у цыплят с недостаточностью витамина B12 оказалась значительно повышенной. Есть основательные данные в пользу того, что новообразованные метильные группы появляются в метионине, но потом в результате трансметилирования они могут оказаться в холине или креатине. Эти выводы никто не оспаривал, но некоторые исследования позволяли предположить, что, кроме того, при недостаточности витамина B12 у крыс активность трансметилазы в печени понижена.
характерного для авитаминозных животных (метод "парного кормления"). Подтверждение данных для поросят и цыплят в опытах с 14С-формиатом и 14С-серином получили Джонсон и сотр. Однако при использовании меченого формальдегида результат оказался неожиданным: интенсивность включения метки в метильные группы метионина и холина у цыплят с недостаточностью витамина B12 оказалась значительно повышенной. Есть основательные данные в пользу того, что новообразованные метильные группы появляются в метионине, но потом в результате трансметилирования они могут оказаться в холине или креатине. Эти выводы никто не оспаривал, но некоторые исследования позволяли предположить, что, кроме того, при недостаточности витамина B12 у крыс активность трансметилазы в печени понижена.
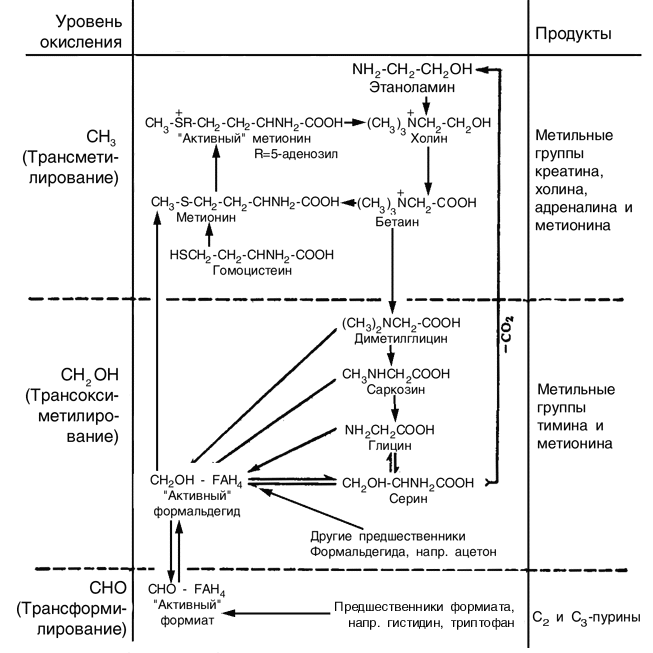
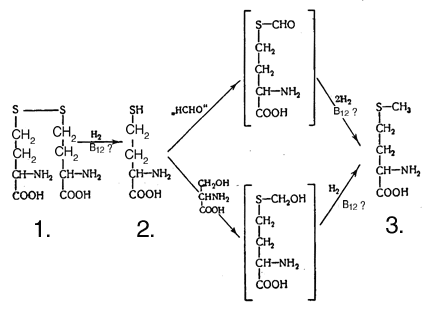
Значение витамина B12 в переносе групп с одним атомом углерода почти неразделимо переплетается с функциями фолиевой кислоты (точнее, производных тетрагидрофолевой кислоты). Эти процессы переноса, которые могут происходить на трех различных уровнях окисления, схематически представлены на схеме 2, показаны также связанные с ними реакции окисления и восстановления. Некоторые из этих процессов переноса происходят в несколько этапов (не показанных на схеме 2) это, безусловно, относится к превращению гомоцистина в метионин, и почти нет сомнений, что для реакции в целом необходимы оба витамина. Возможная последовательность этапов показана на схеме 3.