Бионеорганическая химия
В последнее время выполнены важные работы по моделированию природных систем с металлокомплексами, образованными макроциклическими лигандами. В качестве «модельных» лигандов использовались так называемые «короны» (или краун-эфиры) и «криптаты».
Примером простейших корон, являющихся двухмерными (плоскостными) лигандами, могут служить следующие:

Дициклогексил-14-корона-4 Бензо-18-корона-6
Номенклатура краун-эфиров, как видно из приведенных названий и формул, указывает на общее число атомов в макроцикле и число гетероатомов, формирующих полость короны и выполняющих функции доноров.
Экспериментально установлено, что устойчивость комплексных соединений ионов металлов с коронами определяется соотношением размеров иона металла и полости короны. Наибольшая прочность макроциклических комплексов достигается, когда полость плотно «обхватывает» ион металла. Если полость слишком велика или мала, устойчивость комплексов уменьшается.
Те же закономерности были установлены для систем, в которых комплексообразование ионов металлов осуществляется с помощью макроциклических лигандов - криптатов, представляющих собой трехмерные лиганды с полостью, обрамленной тремя углеродными цепями, включающими гетероатомы. Общая формула криптатов имеет вид:
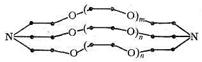
Таким образом, криптаты можно рассматривать как бициклические кислород-донорные лиганды с концевыми атомами, роль которых выполняют третичные атомы азота.
Полагают, что криптаты могут обладать большой избирательностью (селективностью) по отношению к биометаллам, например, природный макроциклический лиганд валиномицин селективен к ионам К+. Ниже приведены значения lgКуст комплексов криптатов и ионов щелочных металлов, образованных лигандами различной дентатности. Число донорных атомов в криптате влияет на размеры полости этого трехмерного макроциклического лиганда и, следовательно, на соотношение размеров комплексообразующего иона металла и полости, а значит и на устойчивость образующегося комплекса:
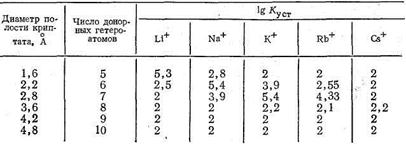
Интересно, что оптимальный лиганд для Li+ содержит в полости макроцикла пять донорных атомов. Увеличение числа донорных атомов и соответственно размеров полости приводит к уменьшению стабильности криптата лития. Для иона Na+ оптимальные размеры полости криптата отвечают 6-дентатному лиганду, для иона К+ - 7-дентатному. Для иона Rb+ устойчивость комплекса в оптимальных условиях ниже, чем для К+, и еще более она падает в случае иона цезия - самого большого по размерам среди ионов щелочных металлов. По-видимому, независимо от размеров полости для большого иона Cs+ прочность связи с лигандом мала из-за уменьшения энергии электростатического взаимодействия Cs+-лиганд.
Итак, исследование криптатов щелочных металлов показывает, что, регулируя состав и геометрию макроциклических лигандов, можно добиться их высокой селективности по отношению к ионам металлов, входящих в состав комплексов.
Состав и строение природных ионофоров, упрощенными моделями которых являются краун-эфиры и криптаты, сложны и многообразны.
Понятно поэтому, что в биосистемах может быть достигнута высокая селективность действия макроциклических лигандов, это и определяет их «узкую специализацию» в процессах метаболизма.
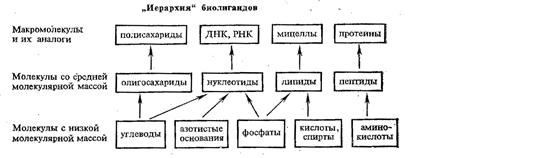
Биополимеры, на основе которых строится комплексообразование в биологических системах
Наиболее важными биополимерами, обеспечивающими процессы обмена веществ в животных и растительных организмах, в том числе процессы, протекающие с участием комплексных соединений металлов, являются полисахариды, белки и нуклеиновые кислоты.
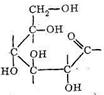
Среди полисахаридов наибольшее значение имеют крахмал, гликоген и целлюлоза. Основным звеном в построении полимерных цепей полисахаридов являются остатки D-глюкозы. Нециклическая D-глюкоза легко циклизуется, образуя две равновесные формы:

При полимеризации α - и β-формы D-глюкопиранозы соединяются в полимерную цепь через кислородные мостики:
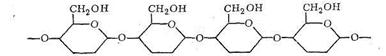
Если полимеризуется α-форма, то цепь полимера оказывается разветвленной - получается крахмал и гликоген. При полимеризации β-формы образуется цепочечный неразветвленный полимер - целлюлоза, которая, как известно, обладает волокнистым строением.
Полисахариды, как видно из приведенных формул, имеют в своем составе кислородные атомы, способные проявлять донорные функции. Таким образом, полисахариды, в виде которых организм запасает углеводы (крахмал, гликоген) и которые используются для построения оболочек растительных клеток (целлюлоза), являются полимерными лигандами.
![]() Другой тип биополимерных лигандов - белки (протеины). Белки представляют собой полимерные образования, в которых в том или ином порядке чередуются 23 α-аминокислоты. Строение всех аминокислот может быть описано формулой
Другой тип биополимерных лигандов - белки (протеины). Белки представляют собой полимерные образования, в которых в том или ином порядке чередуются 23 α-аминокислоты. Строение всех аминокислот может быть описано формулой
Различаются они только природой радикала R. α-Аминокислоты, вступая в реакцию полимеризации, образуют пептидную цепь:
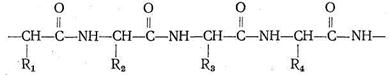
Подобно тому как из 32 букв алфавита путем их различного сочетания можно составить огромное количество слов, так из 23 α-аминокислот посредством их сочленения в том или ином порядке получается все многообразие белковых тел, существующих в природе, образуется так называемая первичная белковая структура. Кроме того, рассматривают вторичную, третичную и четвертичную структуру.
Вторичная структура (α - и β-конформации) возникает в результате взаимодействия полипептидных цепей друг с другом. α-Конформация имеет спиралеобразное строение, каждый виток спирали содержит от трех до семи аминокислотных фрагментов. Взаимодействие между соседними полипептидными цепями в такой спирали осуществляется посредством водородных связей, образованных карбонильным кислородом одной цепи с иминогруппой другой цепи:
![]()
Редко встречающаяся β-конформация содержит вытянутые друг возле друга неспиральные полипептидные цепи.
Третичная структура белка - это глобулы, образованные α-вторичной структурой в результате свертывания полипептидных цепей в клубки. Свертывание α-спиралей в глобулы происходит в результате взаимодействия друг с другом гидрофобных участков спирали, электростатического взаимодействия заряженных участков цепи, образования сульфидных мостиков и водородных связей.
Четвертичная структура возникает в результате объединения глобул в еще более сложную структуру.
Свойства белков как биолигандов определяются содержанием в полипептидных цепях донорных атомов азота и кислорода, которые могут участвовать в образовании хелатных циклов и макроциклических комплексов. Кроме того, к полипептидным цепям через различные функциональные группы могут быть привязаны порфириновые кольца. Порфирин содержит четыре пиррольных ядра (с различными заместителями):