Асимметрия мембран
ВИРУСЫ С ОБОЛОЧКОЙ
Вирусы с оболочкой имеют нуклеокапсид, окруженный липидным бислоем. Последний происходит от мембраны клетки-хозяина и образуется при почковании вируса. Вирус саркомы Рауса и вирус везикулярного стоматита отпочковываются от плазматической мембраны клетки-хозяина в среду, а другие вирусы высвобождаются во внутренние компартменты клетки, например в аппарат Гольджи или эндоплазматический ретикулум. Эти вирусы очень полезны как модели для изучения мембранного биогенеза и внутриклеточного мембранного транспорта. Кроме того, они представляют интерес и с точки зрения изучения образования доменов в мембранах. Как показано на рис. 4.9, процесс отпочковывания включает взаимодействие мембраны с нуклеокапсидом и трансмембранными белками шиловидных выростов вируса. Гликопротеины этих выростов внедряются в плазматическую мембрану клетки-хозяина и с помощью своих цитоплазматических доменов взаимодействуют с белками вирусного матрикса, которые связаны с вирусным нуклеокапсидом. В процессе отпочковывания концентрация белков шиловидных вирусов в растущей почке увеличивается, а белки плазматической мембраны клетки-хозяина полностью из нее исключаются. Сформировавшаяся вирусная оболочка содержит только белки шиловидных выростов и совсем не содержит белков клетки-хозяина. Предполагается, что именно взаимодействие между матриксными белками и белками выростов обусловливает латеральное разделение компонентов, происходящее в плазматической мембране. Интересно, что ли-пидный состав вирусной оболочки не совпадает с составом плазматической мембраны клетки-хозяина. Проще всего объяснить
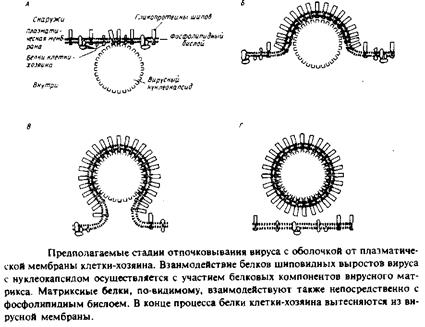
это предпочтительными взаимодействиями между гликопротеинами шиловидных выростов и определенными липидными компонентами. Эта модельная система окажется полезной и для дальнейших исследований латеральной гетерогенности в мембранах.
ЛИПИДНЫЕ МИКРОДОМЕНЫ
При подходящих условиях липиды подвергаются латеральному фазовому разделению с образованием стабильных ламеллярных доменов. Такое разделение можно индуцировать изменением температуры, давления или ионной силы либо добавлением двухвалентных катионов или белков. Вопрос о том, существуют ли микродомены, подобные наблюдаемым в модельных липидных системах, также и в биологических мембранах, всегда вызывал большой интерес У исследователей. Полученные результаты не являются абсолютно убедительными, поскольку липидные домены не удается выделить и охарактеризовать, как в случае латеральной гетерогенности, рассмотренной выше. Конечно, лучше всего было бы провести фрагментацию мембраны и проиллюстрировать различия в распределении компонентов в выделенных мембранных фракциях. Для обнаружения латеральной гетерогенности биологических мембран часто используют электронную микроскопию. О такой гетерогенности можно судить также по данным биофизических методов, если полученный от образца сигнал свидетельствует о наличии различных мембранных популяций, а не одной гомогенной популяции. В качестве примера можно привести измерения коэффициента диффузии флуоресцентных аналогов липидов в протопластах сои. О микрогетерогенности мембран иногда можно судить по поведению ферментов, если ферментативная активность не отвечает усредненному физическому состоянию липидной фазы по всей массе мембраны. Часто эти методы свидетельствуют о наличии в мембране областей с разной текучестью липидов, что указывает на сосуществование фазы геля и жидкокристаллической фазы. Такие выводы были сделаны в ряде исследований, где различные возмущения, вызываемые присутствием цис- и транс-ненасыщенных жирных кислот, объяснялись их разным распределением между доменами, находящимися в жидкокристаллическом состоянии и в фазе геля.
Все эти исследования согласуются с предположением о наличии в биологических мембранах микродоменов, но в большинстве случаев такое объяснение не является единственно возможным или несомненным. Тем не менее идея о существовании липидных микродоменов весьма привлекательна. Ведь благодаря им ферменты в одной и той же мембране могут находиться в разном окружении, и их активность может регулироваться за счет специфичных взаимодействий с липидами или другими белками. Кроме того, на границах между доменами могут проявляться «дефекты» упаковки липидного бислоя. Как полагают, такие границы с резким изменением упаковки липидов в ряде случаев оказывают решающее влияние на функции бислоя. Исследования модельных липидных систем показали, что при фазовом переходе, когда сосуществуют фаза геля и жидкокристаллическая фаза, пассивный транспорт как гидрофобных, так и гидрофильных веществ ускоряется в несколько раз по сравнению с жидкой фазой. По-видимому, в областях бислоя с дефектами упаковки, где сжимаемость липидов высока, могут образовываться поры. Благодаря наличию дефектов в пограничных областях бислоя может ускоряться флип-флоп фосфолипидов, возрастать их доступность для фосфолипаз, а также усиливаться тенденция липидных везикул к слиянию.
Идея липидных микродоменов в мембранах весьма привлекательна, поскольку она позволяет легко объяснить многие факты, но она все еще нуждается в подтверждении.
Резюме
Множество фактов свидетельствует о гетерогенности биологических мембран как в продольном, так и в поперечном направлениях. Трансмембранная асимметрия означает, что разные половины бислоя имеют разный состав. Установлено, что интегральные мембранные белки встроены в мембрану асимметрично и эта асимметрия стабильна. Таким образом, к цитоплазматической и наружной поверхностям мембраны обращены разные белковые домены. Получено множество данных о том, что совершенно разным может быть и фосфолипидный состав двух половин бислоя. Как создается эта липидная асимметрия и за счет чего она поддерживается — пока неясно, хотя имеются данные о существовании АТР-зависимых транслоказ, которые ускоряют перенос липидов через бислой. Другими факторами, определяющими асимметрию мембран животных клеток, являются взаимодействия липидов с цитоскелетом и с внеклеточным матриксом иа ее поверхности.
Имеются данные и о латеральной гетерогенности биологических мембран. Это могут быть достаточно обширные специализированные участки мембраны, например апикальная или базолатеральная области плазматической мембраны поляризованных эпителиальных клеток. Примером того, как в пределах одной мембраны соседствуют области с разным составом и функциями, могут служить менее протяженные соприкасающиеся и несоприкасающиеся участки тила-коидной мембраны. Все это показывает, что молекулярная организация мембран гораздо сложнее, чем это следует из жидкостно-мозаич-ной модели, первоначально предложенной Сингером и Николсоном.