Биологическая функция нуклеиновых кислот
Небольшая молекулярная масса, наличие достаточно большого количества (до 10 %) минорных оснований, которые являются прекрасными маркерами, существенно облегчают проблему определения нуклеотидной последовательности тРНК. В 1965 г. Р. Холли и его сотрудники установили полную нуклеотидную последовательность аланиновой тРНК дрожжей; в 1967 г. А.А. Баев и сотрудники установили последовательность нуклеотидов валиновой тРНК дрожжей. А. Рич и др. (1975—1977 гг.) провели полную расшифровку пространственной структуры фенилаланиновой тРНК на основе рентгенограмм с разрешением до 0,4 нм. Вторичная структура тРНК в плоском изображении имеет вид клеверного листа (рис. 3). тРНК содержит 4 двухцепочечных спиральных участка, 3 из которых являются "шпильками", несущими петли из неспаренных нуклеотидов; 3'- и 5'-концы полинуклеотидной цепи объединены в наиболее длинный спиральный участок, образованный водородными связями между азотистыми основаниями и завершающийся неспаренным тринуклеотидом ССА, Кроме четырех основных ветвей, более длинные тРНК содержат короткую пятую, или дополнительную, ветвь. Две из основных ветвей непосредственно обеспечивают функцию тРНК как адалтора (между двадцатибуквенным кодом белков и четырехбуквенным кодом нуклеиновых кислот). Антикодоновая ветвь имеет антикодон, представляющий собой специфический триплет нуклеотидов, комплементарный кодону мРНК и способный образовывать с ним пары оснований. Акцепторная ветвь присоединяет специфическую аминокислоту за счет образования эфирной связи между ее карбоксильной группой и гидроксильной группой 3'-концевого остатка аденина в тРНК, Две другие главные ветви тРНК называются дигидроуридиловая ветвь и Т![]() С-ветвъ. Первая содержит необычный нуклеозид дигидроуридин, а вторая — нуклеозиды псевдоуридин (
С-ветвъ. Первая содержит необычный нуклеозид дигидроуридин, а вторая — нуклеозиды псевдоуридин (![]() ) и риботимидин (Т), обычно не присутствующие в составе РНК.
) и риботимидин (Т), обычно не присутствующие в составе РНК.
Исследования структуры тРНК методом рентгеноструктурного анализа показали, что их нативные молекулы имеют компактную форму; отдельные двухспиральные "шпильки" клеверного листа складываются в специфическую третичную структуру, которая является близкой для всех тРНК.
После ферментативной этерификации свободной 3'-гидроксигруппы концевого остатка адениловой кислоты в последовательности ССА специфической в отношении тРНК аминокислотой образуется активная форма, называемая аминоацил-тРНК. Остаток этой аминокислоты переносится к концу растущей полипептидной цепи. Антикодон обеспечивает специфичность взаимодействия тРНК с мРНК. Боковые петли, видимо, играют важную роль в связывании тРНК с аминоацил-тРНК-синтетазой и с комплексом рибосома—мРНК. Аддукты аминоцил—тРНК располагаются в определенной последовательности, связанной с последовательностью кодонов мРНК.
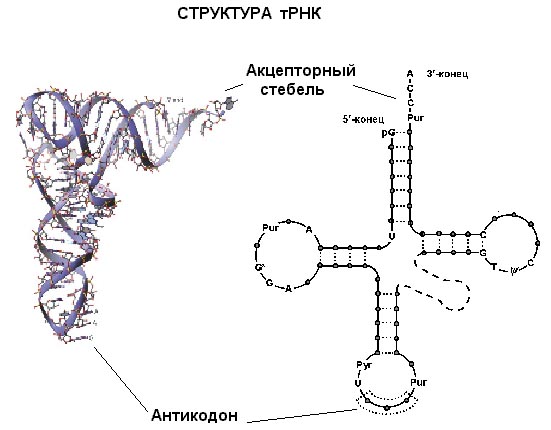
Рис.5 Структура транспортной РНК
Матричная РНК составляет незначительную часть (3—10 %) всех клеточных РНК; молекулярная масса колеблется в широких пределах и доходит до 14 10![]() . Она программирует синтез всех клеточных белков цитоплазмы. Относительное содержание индивидуальной мРНК в суммарном препарате РНК может составлять тысячные доли процента. Первые экспериментальные доказательства существования мРНК получили А.Н. Белозерский, А.С. Спирин и их сотрудники (1957—1960 гг.). Они показали, что нуклеотидный состав общей РНК бактерий E. coli коррелирует с составом их ДНК, и пришли к заключению о наличии, по крайней мере, двух типов РНК, один из которых (большая фракция) имеет состав, не отражающий состава ДНК, а второй (меньшая фракция) воспроизводит состав ДНК. В дальнейшем выяснилось, что первая фракция — это рибосомная РНК, а вторая — мРНК. Но это сделали в 1961 г. Ф. Гросс и сотрудники.
. Она программирует синтез всех клеточных белков цитоплазмы. Относительное содержание индивидуальной мРНК в суммарном препарате РНК может составлять тысячные доли процента. Первые экспериментальные доказательства существования мРНК получили А.Н. Белозерский, А.С. Спирин и их сотрудники (1957—1960 гг.). Они показали, что нуклеотидный состав общей РНК бактерий E. coli коррелирует с составом их ДНК, и пришли к заключению о наличии, по крайней мере, двух типов РНК, один из которых (большая фракция) имеет состав, не отражающий состава ДНК, а второй (меньшая фракция) воспроизводит состав ДНК. В дальнейшем выяснилось, что первая фракция — это рибосомная РНК, а вторая — мРНК. Но это сделали в 1961 г. Ф. Гросс и сотрудники.
Если рРНК и тРНК метаболически устойчивы, то мРНК в большинстве случаев, особенно у прокариот, является относительно короткоживущей. Ее нуклеотидный состав близок к составу ДНК, выделенной из того же организма. мРНК имеют отчетливо выраженную вторичную структуру; в состав двухцепочечных участков включено до 75 % всех нуклеотидных последовательностей мРНК. Значительная часть участков вторичной структуры в мРНК идентифицирована "шпильками". Однако роль участков вторичной структуры в реализации матричных функций пока точно не установлена. Предполагается, что "шпильки" выполняют роль специфических структур, обусловливающих узнавание определенных участков рибосом при их связывании с мРНК.
Если рРНК и тРНК относятся к обслуживающему аппарату белоксинтезирующей системы клетки, то мРНК является прямым посредником между ДНК и белками, играет роль матрицы для синтеза последних, поэтому считают, что она выполняет роль мессенджера. Сама мРНК синтезируется в ядре клетки в процессе транскрипции у в ходе которой нуклеотидная последовательность одной из цепей хромосомной ДНК ферментативным путем "переписывается" (транскрибируется) с образованием предшественника пре-мРНК; последняя имеет копии палиндромов ДНК, поэтому ее вторичная структура содержит шпильки и линейные участки. При созревании пре-мРНК шпильки отсекаются ферментами и образуется мРНК.
2. Материалы и методы исследований
2.1. Кислотный гидролиз нуклеопротеидов дрожжей и изучение свойств ДНК И РНК
Оборудование и реактивы: весы технические; весы торзионные; электроплитка; водяная баня; центрифуга; колба на 100 мл с обратным холодильником; пробирки; пипетки на 1 и 20 мл. Концентрированная и 10%-ная серная кислота; 10%-ный раствор NaOH; концентрированный раствор аммиака; 1% раствор AgNO![]() ; бидистиллированная вода; раствор дифениламина (1 г дифениламина растворяют в 50 мл ледяной уксусной кислоты, добавляют 2,75 мл концентрированной H
; бидистиллированная вода; раствор дифениламина (1 г дифениламина растворяют в 50 мл ледяной уксусной кислоты, добавляют 2,75 мл концентрированной H![]() SO
SO![]() и доводят ледяной уксусной кислотой до 100 мл); молибденовый реактив (18,75 г молибдата аммония растворяют в 250 мл 32%-ного раствора HNO
и доводят ледяной уксусной кислотой до 100 мл); молибденовый реактив (18,75 г молибдата аммония растворяют в 250 мл 32%-ного раствора HNO![]() ); 1%-ный раствор CuSO
); 1%-ный раствор CuSO![]() ; 1%-ный раствор тимола.
; 1%-ный раствор тимола.
Материалы: дрожжи сухие; препараты ДНК и РНК.
Ход работы
1. Гидролиз нуклеопротеидов. В коническую колбу на 100 мл вносят 1 г сухих дрожжей, добавляют 20 мл 10%-ного раствора H![]() SO
SO![]() и 20 мл бидистиллированной воды. Колбу соединяют с обратным холодильником, нагревают до кипения и кипятят в течение 1 ч, охлаждают и центрифугируют 5 мин при 5000 об/мин.
и 20 мл бидистиллированной воды. Колбу соединяют с обратным холодильником, нагревают до кипения и кипятят в течение 1 ч, охлаждают и центрифугируют 5 мин при 5000 об/мин.
2. Качественные реакции на пентозы:
а) к 0,5 мл нейтрализованного щелочью гидролизата добавляют 2 капли 1%-ного спиртового раствора тимола, перемешивают и осторожно прослаивают равный объем концентрированной серной кислоты. На дне пробирки образуется красное окрашивание в результате конденсации тимола с фурфуролом, получившимся из пентозы;