Возможное адаптивное значение открытого таза птиц и новая гипотеза происхождения полета
Таким образом, если эволюционное исчезновение симфизов лобковой и седалищной костей можно связать с крупными яйцами, то разворот их назад как будто даже противоречит этому.
Роль своеобразного таза птиц в дыхании, учитывая существенные отличия дыхательного акта птиц и млекопитающих, невозможно оценить без сложных физиологических методик. Особенности дыхания вымерших птицеподобных форм динозавров нам неизвестны, и поэтому мы не беремся подтверждать или опровергать данную версию.
Что касается бипедализма, т. е. передвижения преимущественно или исключительно на задних конечностях, то оно достаточно широко распространено среди современных и вымерших форм млекопитающих (Соколов, 1973) и рептилий (Коуэн, 1982; Кэрролл, 1993). Ни у кого из млекопитающих, как прыгающих на задних ногах с синхронным и асинхронным отталкиванием (кенгуру, тушканчики), так и способных к шаговому передвижению на задних конечностях (некоторые виды тушканчиков) не наблюдается тенденции к изменению формы таза. К непродолжительному бегу на задних конечностях способны некоторые современные ящерицы — василиски (Basiliscus sp.), плащеносная ящерица (Chlamydosaurus kingi), однако и у них таз типично трехлучевого рептильного типа. В то же время у современных крокодилов, несмотря на препубический таз (с направленными вперед лобковыми костями) отсутствует лобковый симфиз (Шимкевич, 1922). Среди вымерших групп рептилий обращают на себя внимание формы, относимые к двум группам динозавров — ящеротазовых (Saurisсhia) и птицетазовых (Ornithisсhia), первые из которых имели типично рептильный таз с направленными вперед и вниз лобковыми костями, образующими в районе симфиза массивное утолщение — “башмак”, хотя у некоторых из них, в первую очередь у двуногих, происходило расширение подвздошной кости и прирастание ее к значительному количеству позвонков. У второй группы подвздошные и лобковые кости отогнуты назад, как у птиц. У некоторых форм подвздошная кость срастается с немногими (4–6) позвонками, у других расширяется и охватывает большее число позвонков. У ряда специализированных форм (особенно более крупных поздних — игуанодонтов, стегозавров) как двуногих, так и четвероногих, развивается направленный вперед предлобковый отросток, который занимает положение, сходное с лобковой костью ящеротазовых. И среди первых, и среди вторых описаны формы, передвигающиеся исключительно в двуногом положении. Некоторые из них, по-видимому, были способны к быстрому и маневренному бегу (Рауп, Стенли, 1974; Коуэн, 1982). Положение отдельных частей позвоночного столба и грудной клетки было сходным с птичьим. Сходством обладала длинная и подвижная шея и легкий ажурный череп, у ряда форм челюсти, по-видимому, одевал клюв (Кэрролл, 1993). Сходство ряда ящеротазовых форм с бегающими птицами было настолько велико, что они получили название Ornithomimidae и Avimimidae. Особенно высокое сходство отмечено в скелете задних конечностей, что вероятнее всего обусловливало и сходство мускулатуры. В то же время среди птицетазовых, несмотря на меньшее общее сходство, встречались, особенно среди “примитивных” ранних, формы, весьма сходные с птицами по строению задних конечностей (Heterodontosaurus, Hypsilophodon), таза (Scelidosaurus, Protoceratops, Homalocephale), часть из которых передвигалась преимущественно или исключительно на задних конечностях, часть использовала исключительно четвероногое передвижение. Несмотря на ряд попыток объяснения подобных различий в строении таза динозавров (Romer, 1956; Galton, 1969; Charig, 1972; Walker, 1977; Santa Luca, 1980, цит. по: Кэрролл, 1993), большинство ученых так или иначе связывают их с локомоцией и реже с объемом брюшной полости (кишечника). Единого мнения так и не сложилось. Однако наличие сходных по габитусу и локомоции форм с разным строением таза само по себе опровергает такую связь (рис. 1). Существенным отличием, правда, всех этих форм, кроме, видимо, Avimimus, является более вертикальное положение конечностей, чем у современных птиц, особенно направленное вниз, а не вперед, бедро, а также значительно меньшая ширина таза, связанная с направлением хотя и широких подвздошных костей косо вниз, а не в стороны, как у птиц, что создавало все-таки заметно меньший объем тазовой полости. Таким образом, связывать характерное положение лобковых и седалищных костей птиц с двуногостью видимо все-таки нет оснований. Как хорошо показал Курзанов (1987), смещению лобковой и седалищной костей сначала вниз, а потом назад не только у птиц, но и у птицеподобных динозавров предшествовало изменение положения бедра и выход его в нормальном (при стоянии) положении вперед от лобковой кости. Это заставило бегающих авимимид изменить расположение и места прикрепления мускулатуры, связанной с бедром, и освободило в значительной степени, а возможно и полностью, лобковую кость от функции прикрепления мускулатуры, связанной с бедром и способствовало дальнейшей возможности изменения ее положения. По-видимому, такая же ситуация сложилась при развитии ранних бегающих предков птиц, хотя палеонтологические остатки, на которых можно было бы показать этот процесс, отсутствуют. Обоснований подобного поворота бедра несколько, но нам кажется, что наиболее существенное — в изменении положения центра тяжести при статической позе, который смещается вперед при уменьшении массы либо укорочении хвоста. С данной проблемой столкнулись как авимимиды, так и птицы (но не орнитомимиды и другие бегающие динозавры с развитым хвостом).
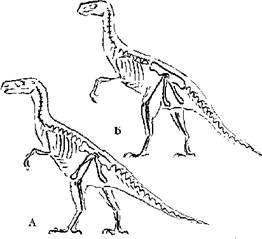
Рис. 1. Строение скелета ящеротазового (А) и птицетазового (Б) динозавров.
Еще меньше вероятность связи птичьего таза с полетом, так как у летающих птерозавров (Pterosauria) таз имел обычное строение (см. также летучие мыши).
Предлагаемое нами объяснение строения таза
Таким образом, рассмотрев предыдущие гипотезы, мы видим, что они не могут быть во всяком случае ведущими в становлении своеобразного для птиц (и ряда динозавров) строения таза. В то же время, вскрывая 1–3-суточных птенцов выводковых птиц, можно обнаружить высокую степень приспособленности данного строения таза с широкими, почти горизонтально поставленными подвздошными костями, широко разнесенными вертлужными впадинами и отодвинутыми назад вверх лобковыми и седалищными костями к вмещению огромного запаса желтка (рис. 2, 3). Этот запас желтка способен поддержать существование птенца иногда до недельного возраста (при оптимальной температуре) без питания (Хейнрот, 1947). Подобный запас желтка, более или менее выраженный, имеют все выводковые птицы. У птенцовых (воробьиные, голуби) он намного меньше, даже у только что вылупившихся птенцов и составляет менее 5 % веса тела (Дементьев, 1940; Хейнрот, 1947; Флинт и др., 1986). Эти различия отражают две экологически разные стратегии эволюции птиц. Типичных птенцовых родители обогревают постоянно и с первых же часов начинают кормить. Запас желтка в такой ситуации практически не нужен, и почти весь желток, находящийся в яйце, еще на ранних стадиях развития идет на построение тела зародыша. Соотношение желтка к белку в яйцах таких птиц 1:2,5 — 1:4 (голуби), 1:3 (дрозды, ласточки), 1:4 (домовый воробей (Passer domesticus)) (Дементьев, 1940). У видов же выводковых соотношение от 1:2 до 1:0,5. При этом часть желтка расходуется в первой трети эмбрионального развития (зародышевый период) на построение тела, а остальная часть (более 2/3) не используется во время эмбрионального роста, когда в предплодном и плодном периоде зародышевого развития эмбрион строит свое тело за счет белка яйца и незадолго перед вылуплением втягивается в брюшную полость, создавая запас на первые дни жизни (Дементьев, 1940; Флинт и др., 1986). Этот запас особенно велик у современных форм, забота родителей о вылупившихся птенцах у которых отсутствует и птенцам приходится самостоятельно обучаться питанию, защитным реакциям и поддерживать температурный баланс. Речь идет о сорных курах (Megapodus eremita, Leipoa ocellata, Telegallus sp.). Соотношение желтка к белку у этих птиц 1:0,5; 1:0,66 соответственно (Хейнрот, 1947). У видов, самостоятельно питающихся с первого дня жизни и находящихся в достаточно жестких температурних условиях, соотношение ниже, например, у гуся — 1:0,9–1:0,97. У видов, первые дни которых проходят в более оптимальных условиях и родители их могут помогать кормодобыванию, соотношение еще ниже: цесарка (Numida meleagris) — 1:1,2; индюк (Meleagris galloopavo), домашняя утка (Anas platyrhynchos) —1:1,4; коростель (Crex crex) — 1:1,6. У курицы (Gallus domesticus), как одомашненного вида, соотношение варьирует — 1:1,7–1:2. У современных птиц в силу широкой адаптивной радиации встречаются все сочетания черт “выводковости” и “птенцовости” вплоть до характерного для чаек и крачек рождения зрячих опушенных и способных к некоторой терморегуляции птенцов, которых все же выкармливают родители до подъема на крыло. Тем не менее, общепринятым считается, что выводковый тип развития для птиц исходен. Наиболее критическим в таких случаях является первый период после вылупления, за который птенцы должны освоить способы поиска и потребления корма и защиты от опасности, хотя в этом им могут помогать родители. При птенцовом типе развития этот критический период исчезает и вся энергия, потребляемая птенцами с кормом, идет на рост тела. У птенцовых развитие и рост идут быстрее. У сходных по размерам и весу сизого голубя (Columbia livia) и серой куропатки (Perdix perdix), стартующих с примерно равных весовых значений, окончательный вес и размер достигается соответственно к 30–40 и 100–120 дню (Поярков, 1955). У наиболее мелких выводковых этот период не меньше 5 недель, у крупных — до 6 месяцев, в то время как достижение окончательных линейных размеров даже у самых крупных птенцовых не более 2 месяцев. У большинства птенцовых более длинный период занимает лишь развитие крупных партий полетного пера (Дементьев, 1940; Чельцов-Бебутов, 1982).