Возможное адаптивное значение открытого таза птиц и новая гипотеза происхождения полета
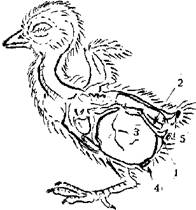
Рис. 2. Размещение запаса желтка у суточного цыпленка:
1 — лобковая кость pubis;
2 — седалищная кость pinbone;
3 — втянутый желток drawn yolk;
4 — пупочное кольцо navel ring;
5 — отверстие клоаки cloacal foramen.
Аналогичный птичьему запас желтка имеется при вылуплении в яйцах черепах и крокодилов, и этот запас тоже расходуется в критический ранний период до перехода на автономное питание. У крокодилов их может хватить иногда на месяц (Даревский, Орлов, 1988), а у черепах, возможно, и на больший срок, но это связано с пойкилотермностью данных видов. У змей и ящериц относительный вес желтка в яйце тоже велик, однако это связано, по-видимому, с тем, что в плодный период развития белок слабо используется для построения тела животного. Запасы желтка при рождении у этих групп незначительны. Высокая постоянная температура тела птиц создает дополнительные сложности и сокращает период, обеспечиваемый питательными веществами втянутого желточного мешка и приводит к необходимости увеличения его объема. У современных крокодилов и черепах при их пропорциях тела как взрослых, так и молоди, не создавалось особых трудностей при помещении значительных объемов остаточного желтка (у черепах внутри панциря, у крокодилов между последними ребрами и отодвинутым назад тазовым поясом). У современных птиц, как и у их ранних форм (насколько мы можем судить по археоптериксу (Archaeopterix lithographica) при коротком туловищном отделе размещение значительных объемов желтка становится сложным, особенно после развития массивной грудины, простирающейся далеко назад. Единственным способом было отодвинуть назад и в стороны элементы тазового пояса. Казалось бы, эта проблема раннего постэмбрионального периода не должна существенно влиять на морфологию взрослых форм, но для птиц как класса в целом одной из характернейших особенностей является детерминированный рост. Пожалуй не найдется ни одной другой группы позвоночных, имеющей такое соотношение размеров новорожденных и взрослых форм (близкое соотношение можно найти только у тоже летающих летучих мышей).
Практически все птицы начинают пользоваться полетом только после достижения линейных размеров взрослой формы. Несколько затушевывают эту важнейшую закономерность несовершенная система промеров птиц, где один из основных — длина крыла с учетом маховых перьев и использование для определения размеров весовых показателей, которые у молодых птиц обычно ниже, чем у взрослых. Хотя есть ряд видов и даже групп, у которых птенцы перед первыми в своей жизни полетами весят больше взрослых за счет больших запасов жира — трубконосые, стрижи, ласточки (Чельцов-Бебутов, 1982). И несколько выпадают из общей закономерности куриные птицы, у которых в связи с особенностями возрастного развития оперения способными к короткому полету оказываются особи, значительно не достигшие окончательных размеров (Хейнрот, 1947). Все остальные виды к началу этапа активного полета оказываются с полностью сросшимися элементами костей, особенно костей крыла, а в большинстве случаев и со значительно пневматизированными участками скелета (Дементьев, 1940; Дольник, 1975; Флинт и др., 1986). По-видимому, продолжение линейного роста и полет — вещи в эволюционном плане несовместимые. Об этом говорят, во-первых, отсутствие линейного роста после начала полета у рукокрылых (Бобринский и др., 1965; Соколов, 1973; Мосияш, 1985) и отсутствие следов линейного роста на костных остатках птерозавров (Кэрролл, 1993).
Второй особенностью птиц как класса является лимитирование у них максимальных, к тому же не очень крупных размеров. Уже давно показана невозможность активного полета у птиц весом более 10–12 кг (Дементьев, 1940; Гладков, 1949; Кокшайский, 1966; Шмидт-Ниельсен, 1978). Мало того, более поздними работами показано, что оптимальный вес для активно летающей птицы лежит в пределах 0,1–1 кг (Кокшайский, 1970; Шмидт-Ниельсен, 1987), а возможно даже в пределах 0,1–0,5 кг (Цвелых, 1983), поскольку более крупным летающим формам приходится формировать специальные морфологические приспособления (увеличение площади крыла, разрезные устройства конца крыла) для покрытия дефицита мощности полетной мускулатуры (Гладков, 1949; Кокшайский, 1966; Якоби, 1966). Птицы весом более килограмма либо редко и несистематически пользуются полетом (дрофы, журавли, куриные), либо переходят к различным типам планирующего полета (трубконосые, аисты, хищники-падальщики), либо полет занимает в их бюджете времени крайне незначительную долю, поскольку энергозатраты на него у таких видов непропорционально велики (Дольник, 1982; Чельцов-Бебутов, 1982). Данное утверждение не касается периода миграций, в течение которого птицы расходуют запасенный в значительных количествах жир и таким образом имеют отрицательный энергобаланс (Дольник, 1975). В целом большинство крупных видов современных птиц относится к высокоспециализированным формам. С другой стороны, наиболее примитивные ранние птицы не могли быть слишком мелкими (типа современных воробьиных) т. е. менее 100 г, т. к. таким формам при полете сложнее преодолевать возникающие завихрения воздуха и ветер (Гладков, 1949; Якоби, 1966; Цвелых, 1983), но, что более важно, у таких мелких современных птиц тоже очень напряженный энергобаланс в связи с противоречием между постоянно поддерживаемой высокой температурой тела и малыми размерами, при которых сохранение тепла превращается в сложную проблему (Шмидт-Ниельсен, 1987; Межжерин, 1988). Таким образом, ранние формы птиц, еще плохо летающие, со слабой полетной мускулатурой, не могли быть слишком крупными, но они не могли быть и слишком мелкими в связи с низкой энерговооруженностью (метаболизм ближе к рептильному, чем к птичьему типу) и, возможно, менее совершенными термоизолирующими качествами покровов.
Следующим существенным моментом для птиц являлась необходимость как можно большего сокращения периода зависимости от родителей (во время гнездования птицы “привязаны” к определенному месту), т. к. до достижения окончательных размеров птицы не могли пользоваться полетом. Все это привело к формированию у птиц, пожалуй, максимально известных среди позвоночных относительных скоростей роста (Шмальгаузен, 1938; Дементьев, 1940; Хейнрот, 1947; Межжерин, 1988). Так, у дроздов, жаворонков, славок линейный рост костных элементов заканчивается иногда к 6–9 дню жизни, и даже у средних по размерам голубей и врановых линейный рост (не учитывая оперения) прекращается к 15–20 дню жизни. Вторым способом сократить период гнездования было увеличение “стартовых значений” для постэмбрионального роста, т. е. увеличение объема и энергетических запасов яйца. Из этого противоречия (крупные размеры яйца и вылупляющегося эмбриона и небольшие окончательные размеры взрослой особи, не меняющиеся с возрастом) сформировалась ситуация, при которой не было времени и “запаса” линейных размеров, за счет которых изменялась бы существенно морфологическая структура, характерная для ранних этапов. В некоторых случаях современные птицы при вылуплении имеют линейные показатели ряда частей тела, характерные для взрослых птиц (лапа некоторых чаек (Larus minutus) и крачек (Sterna hirundo, S. albifrons, род Сhlidonias), цевка ряда мелких чистиковых (Alle, Cepphus, Aethia). Кроме того, увеличенное пространство брюшной — тазовой — полости после использования эмбрионального желтка занималась разросшимся желудком и кишечным трактом у подрастающих особей, а у взрослых позволяла без особого напряжения и “ущемления прав” желудочно-кишечного тракта формировать в яичнике во время размножения крупные яйцеклетки, поскольку при размножении у птицы накладываются друг на друга процессы быстрого роста яйцеклеток в яичнике, формирование белковой и скорлуповой оболочки у находящихся в нижнем отделе яйцевода яиц, причем эти процессы возможны только при интенсивном поступлении питательных веществ и энергии, т. е. опять-таки при максимально эффективной работе пищеварительного аппарата (Дементьев, 1940; Дольник, Дольник, 1982; Флинт и др., 1986). Хорошо известна зависимость яйценоскости, объема яиц, качества желтка и скорлупы и даже ее окраски от интенсивного и сбалансированного питания. Так, например, третьи яйца в кладках чайковых, четвертые — у куликов (группы с детерминированным количеством яиц в кладке) и последние яйца в гнездах воробьиных бледнее и тусклее окрашены, вплоть до совсем белых, и часто мельче по размеру. В то же время, большинство современных рептилий в период роста и формирования яиц, а также на период размножения ограничивают себя в питании либо полностью прекращают питаться, используя накопленные заранее питательные вещества. Крупные размеры эмбрионов у птиц привели к более сложному и дифференцированному развитию в период инкубации и к смещению ряда морфологических изменений на этот период. Так, формирование типичного птичьего таза из возникающей вначале трехлучевой, сходной с рептильной, закладки происходит у куриного, например, эмбриона на восьмой день (Шимкевич, 1922). Втягивание же желтка, как весьма вязкой структуры, происходит в течение нескольких последних дней инкубации и обычно идет параллельно уже начавшемуся легочному дыханию. При этом пупочное отверстие, окруженное кольцевой мускулатурой, имеет значительный диаметр. На этом этапе инкубации довольно велик процент гибели эмбрионов, кроме того, у части их так и не происходит полного втягивания, и пупочное отверстие не закрывается (рис. 3). Такие птенцы, даже нормально вылупившись, без дополнительных хирургических операций обречены на гибель. Таким образом, становится понятным своеобразие темпов роста некоторых структур в постэмбриональном развитии птиц.