Индивидуальное развитие как новая стратегия эволюции
Живые организмы, появившиеся 3–4 миллиарда лет назад в условиях Праземли, были еще весьма примитивны. "Протоклетка" была своего рода микрореактором, содержавшим в растворе полинуклеотиды и полипептиды, а также другие вещества. Этот реактор был отделен от окружающей среды мембраной и одновременно связан с окружающей средой путем обмена водой и растворенными в ней веществами.
Протоклетки обладали способностью к самовоспроизводству с небольшими ошибками, конкурировали друг с другом и были подвержены давлению отбора. В силу уникальных физико-химических свойств полинуклеотидов протоклетки были наделены способностью к хранению и передаче информации, закодированной в последовательности оснований. Новые протоклетки возникали в результате деления клеток. Дочерние клетки после непродолжительной физико-химической фазы роста становились неотличимыми от материнских клеток по всем свойствам, дочерние клетки не претерпевали индивидуального развития и по крайней мере в принципе были бессмертны. Многие из ныне, живущих простейших микроорганизмов, например, вирусы и бактерии, лишены индивидуального развития, они "рождаются в готовом виде" и живут столько, сколько позволяют условия окружающей среды. У более сложных одноклеточных организмов, например, у дрожжевых клеток, наблюдаются отчетливо выраженные жизненные циклы с фазами юности, зрелости, размножения и старости. С возникновением многоклеточных организмов индивидуальное развитие занимает все более важное место.
Наиболее ярким примером может служить человек. Между грудным ребенком и зрелым взрослым человеком имеются существенные качественные различия не только с точки зрения морфологии. Младенец и взрослый отличаются и по своей работоспособности, и по своим интеллектуальным возможностям.
На ранней фазе эволюции живого первые многоклеточные организмы появились лишь как результат случайного образования кластеров. По своим свойствам они еще несущественно отличались от одноклеточных организмов. Индивидуальное развитие как новая стратегия складывалось лишь постепенно, и последующий успех этой стратегии опирался на ее значительные эволюционные преимущества. Онтогенез, характеризуемый как процесс индивидуального развития, является большим достижением эволюции, типичным лишь для более поздних стадий филогенеза – исторического развития таксономических групп. Онтогенез – непременная предпосылка не только образования более сложных морфологических структур, но и процессов обучения, на которых основывается работоспособность всех высших организмов. Поскольку ген вследствие ограниченной емкости его "памяти" может передавать только информацию, важную для выживания, основной поток информации осуществляется через процессы обучения. Возможность обучения является важным преимуществом при отборе. Процессы обучения открывают совершенно новые возможности обработки информации. Поскольку онтогенез занимает в эволюции центральное положение, его эволюционное моделирование имеет большое значение.
2. Дискретные модели циклов жизни
Важные свойства систем, элементы которых претерпевают индивидуальное развитие, могут быть изучены даже на простых дискретных моделях. В простейшем случае жизненный цикл состоит только из двух стадий – молодость и зрелость, которые мы обозначим номерами 1 и 2.
Рассмотрим в качестве примера популяцию клеток прокариотов, например, бактерий. Индивидуальное развитие клеток прокариотов можно разделить на две стадии.
1) Молодость клетки; на этой стадии, продолжающейся около 40 минут, делится ДНК клетки.
2) Зрелость клетки.
Развитие популяций эукариотов, например, дрожжевых клеток, может быть разделено на следующие фазы.
1) Молодость клетки: клетка сосредоточивается на синтезе белка.
2) Зрелость клетки: синтез ДНК; после различных дальнейших этапов, наконец, наступает
3) Митоз.
Если ограничиться двумя стадиями, то процесс моделируется следующими уравнениями;
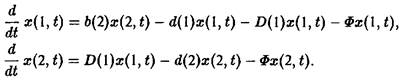
Здесь 6 характеризует размножение, а также порождение молодых клеток зрелыми клетками, d – смертность и D – переход молодость–зрелость.
Величина Ф означает разбавление потока для создания давления отбора. В простейшем случае неумирающих клеток имеем:
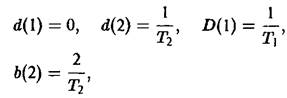
где Г, – продолжительность i-й фазы.
Скорости размножения могут в определенных пределах замедляться. Такое торможение регулируется производством чалонов – ингибиторов клеточного деления. Это приводит к зависимости скорости роста от концентрации. Если ингибиторы испускаются только на стадии зрелости, то скорость роста можно описать следующим выражением:
![]()
В этом случае в системе существует и колебательный режим. Если же ингибиторы производятся главным образом на стадии молодости клетки, то скорость роста описывается следующим выражением:
![]()
где п – надлежащим образом выбранный показатель.
Конкуренция и отбор могут быть введены следующим образом. Рассмотрим систему, состоящую из различных сортов, в две стадии:
![]()
и введем конкуренцию с помощью зависящего от времени разбавления потока:
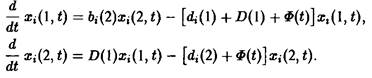
При условии постоянного общего числа частиц получаем:
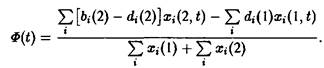
Вторая возможность состоит в том, чтобы создать давление конкуренции посредством связи через субстрат, т.е.
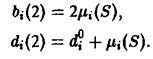
Здесь ![]() – естественная смертность зрелых клеток,
– естественная смертность зрелых клеток,![]() – скорость деления в зависимости от концентрации субстрата. Для описания расхода субстрата используется дополнительное уравнение:
– скорость деления в зависимости от концентрации субстрата. Для описания расхода субстрата используется дополнительное уравнение:
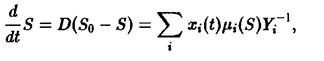
где Y-1i – мера эффективности использования субстрата.
Более подробное обсуждение моделей с двумя и более стадиями индивидуального развития, а также их приложений можно найти в литературе.
3. Модель старения Маккендрика фон Фёрстера
Для математического описания индивидуального развития Маккендрик и фон Фёрстер разработали рекуррентную модель, в которой непрерывный процесс старения рассматривается как последовательность отдельных, изолированных, актов. С математической точки зрения речь идет о том, чтобы вывести систему дифференциальных уравнений для функции плотности x, описывающей, сколько особей возраста г существуют в системе в момент времени t. Рассмотрим такую систему
в момент времени t + At. Число особей, которые к этому времени достигнут возраста г, равно числу особей, которые к моменту времени t достигли возраста г – At, за вычетом особей, умерших за интервал времени At. В результате мы получаем: