Исследование соотношения в мышцах С- и Х-белков в норме и при патологии
Другой амилоидный краситель Конго красный также специфически взаимодействует с амилоидными фибриллами (Klunk et al., 1989). Конго красный – сульфонированый азокраситель с гидрофобной центральной частью, состоящей из бифенильной группы, расположенной между отрицательно заряженными концами молекулы красителя (рис. 8). При связывании Конго красного с амилоидными фибриллами наблюдается зеленое двойное лучепреломление в поляризационном микроскопе, и это свойство делает Конго красный наиболее используемым красителем для диагностики амилоидов. В спектральных исследованиях регистрируется сдвиг спектра поглощения Конго красного в состоянии, связанном с амилоидными фибриллами, в длинноволновую область спектра, а именно от ~490 нм к ~500 нм.
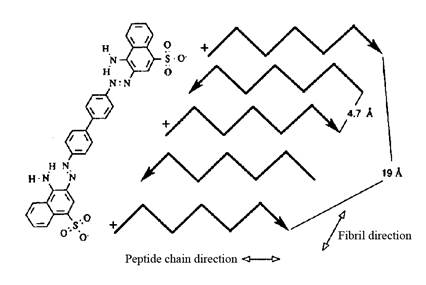
Рис. 8. Модель связывания Конго красного с амилоидными фибриллами. Показан антипараллельный слой, где каждая пятая цепочка белка имеет одинаковое N–C направление. Поэтому, молекула Конго красного может связываться с таким же типом аминокислоты в обеих полипептидных цепях (первой и пятой, как указано на рисунке). Следующая молекула красителя сможет связаться с третьей и седьмой цепочкой и т.д. (Klunk et al., 1989).
Связывание амилоидных фибрилл с Конго красным зависит от структуры амилоидных фибрилл, а именно от наличия β-складчатой структуры с отдельными β-слоями. Кланк и соавторы предложили модель связывания амилоидных фибрилл с Конго красным посредством связей между двумя отрицательно заряженными сульфоновыми группами Конго красного и двумя положительно заряженными аминокислотными остатками двух отдельных белковых молекул, которые определенным образом ориентированы в β-складчатой структуре фибрилл, образованной посредством бок о бок расположенных отдельных молекул (Klunk et al., 1989). Это означает, что белковые цепочки расположены на расстоянии 4.7 Å. Более того, каждая пятая цепочка расположена от первой на расстоянии 19 Å. Это, приблизительно, соответствует расстоянию между сульфоновыми группами КК (рис. 8). Данная модель показывает специфичность взаимодействия Конго красного с амилоидными фибриллами.
2.5. Патологические проявления амилоидозов
К настоящему времени выделяют следующие формы амилоидозов (Мягкова, 2000):
1) первичный (идиопатический) амилоидоз – развивается вследствие невыясненных причин;
2) вторичный (приобретенный) амилоидоз – развивается как осложнение после хронических заболеваний, при которых происходит распад тканей (туберкулез, бронхоэктатическая болезнь, хронический остеомиелит и др.);
3) наследственный (генетический, семейный) амилоидоз – врожденное нарушение белкового обмена;
4) старческий амилоидоз.
С другой стороны, амилоидоз разделяют на системный и локальный. Однако классификация системного амилоидоза основана на специфичности основного амилоидного белка.
Внешний вид органов при амилоидозе зависит от степени развития процесса. Если амилоидные отложения небольшие, внешний вид органа изменяется мало и амилоидоз обнаруживается лишь при гистологическом исследовании. При выраженном амилоидозе орган увеличивается в объеме, становиться очень плотным и ломким, а на срезе имеет своеобразный восковидный или сальный вид.
До сих пор остается нерешенным вопрос, касающийся механизма действия амилоидных фибрилл на органы и ткани. Исследования последних лет показали, что амилоидные агрегаты разных белков и пептидов вызывают нарушение жизнедеятельности клеток и их гибель. Как оказалось, цитотоксические свойства проявляют все амилоидные белки (Hashimoto et al., 2003; Qahwash et al., 2003; Sirangelo et al., 2004; Lee et al., 2006), однако неизвестно, чем обусловлена цитотоксичность этих агрегатов – механическим повреждением клеток, связанным с накоплением амилоидных отложений или особым молекулярным механизмом их взаимодействий внутри клетки.
цель исследования
Выяснение способности саркомерных белков семейства тайтина формировать амилоидные фибриллы in vitro.
задачи исследования
1. Изучить агрегационные свойства саркомерных белков семейства тайтина (тайтин, Х-, С- и Н-белки) in vitro.
2. Изучить агрегационные свойства Аβ(25-35)-пептида в сравнении с агрегацией молекул Х-белка.
3. Проверить амилоидную природу агрегатов, образуемых исследуемыми белками, поляризационной, флуоресцентной микроскопией и спектральными методами.
4. Изучить скорость образования амилоидных фибрилл.
Глава 3. Материалы и методы исследования
3.1. Экспериментальный и клинический материал
Экспериментальный материал: скелетные мышцы и миокард кролика.
Клинический материал: образцы миокарда пациента при ДКМП, взятые при проведении операции по пересадке сердца. Образцы миокарда человека были предоставлены ФГУ НИИ трансплантологии и искусственных органов Росздрава (г. Москва).
3.2. Выделение и очистка белковых препаратов
3.2.1. Очистка С-белка, Х-белка и Н-белка
Х-белок, С-белок и Н-белок очищали по методу (Offer et al., 1973). Фракции Х-, С- и Н-белков, полученные при хроматографической очистке миозина скелетных мышц на колонке с носителем DEAE-Sephadex А-50, концентрировали сульфатом аммония до степени насыщения 2.08 М и осаждали центрифугированием в течение 1 часа при 3000 g. Осадок растворяли в буфере, содержащем 0.3 M KCl, 4.8 мM K2HPO4, 5.2 мM KH2PO4, 0.1 мM ДТТ, 0.1 мM NaN3, pH 7.0, и диализовали против этого буфера до полного удаления сульфата аммония. Разделение белков проводили на колонке с гидроксиапатитом, уравновешенным в этом же буфере, для снятия белков с колонки использовали фосфатный градиент. Такая же процедура очистки применялась и для С-белка миокарда кролика и человека.
3.2.2. Выделение тайтина из скелетных мышц
Тайтин из скелетных мышц кролика выделяли по методу (Soteriou et al., 1993) с модификациями. Мышцы гомогенизировали в 3-х кратном (по отношению к массе мышц) объеме раствора, содержащего 50 мМ KCl, 5 мM ЭГТА, 1 мM NaHCO3,, 1 мM ДТТ, 0.1 мM NaN3, pH 7.0. Для уменьшения деградации тайтина в процессе выделения в раствор добавляли набор ингибиторов протеаз: 1 мM PMSF, 20 мкг/мл ингибитора трипсина, 10 мкг/мл леупептина.
Полученный мышечный гомогенат центрифугировали в течение 5–10 минут при 2500 g. Супернатант отбрасывали, а процедуру промывки повторяли 5–6 раз. К промытому осадку добавляли 2-х кратный объем экстрагирующего раствора, содержащего 0.9 М KCl, 2 мM MgCl2, 2 мM ЭГТА, 0.5 мM ДТТ, 0.1 мM NaN3, 1 мM PMSF, 10 мM имидазола-HCl, pH 7.0. Раствор также содержал 40 мкг/мл соевого ингибитора трипсина и 20 мкг/мл леупептина.
Экстракция проводилась на льду при 4˚С в течение 10–15 минут при непрерывном перемешивании. Экстракт осветляли в течение 40 минут при 14000 g и супернатант разбавляли в 3 раза охлажденной бидистиллированной водой, содержащей 0.1 мM ДТТ и 0.1 мM NaN3, для преципитации актомиозина (конечная ионная сила ~0.2). Через 1 час супернатант осветляли в течение 60 минут при 20000g. Для осаждения тайтина супернатант разбавляли в 5 раз (конечная ионная сила ~0.05) охлажденной бидистиллированной водой, содержащей 0.1 мM ДТТ и 0.1 мM NaN3. Через 40–60 минут осадок, содержащий преимущественно тайтин, собирали центрифугированием в течение 30 минут при 15000 g. Осадок растворяли в минимальном объеме буфера, содержащего 0.6 М KCl, 30 мM KH2PO4, 1 мM ЭГТА, 0.1 мM ДТТ, 0.1 мM NaN3, pH 7.0, и осветляли в течение 60 минут при 20000 g. Тайтин очищали методом гель-фильтрации на колонке с носителем Sepharose–CL2B, уравновешенным в этом же буфере.