Источники и пути образования оксида азота в организме
NOи нервная система
В нервной системе NO имеет большое значение, как в нормальных физиологических условиях, так и при различной патологии. Источниками NO в ЦНС являются нейроны, нейроглиальные клетки – астроциты и клетки микроглии и эндотелий кровеносных сосудов [2].
Нейроны, содержащие NO-синтаза, находятся во многих отделах ЦНС и большинстве изученных периферических ганглиев нервной системы. В коре больших полушарий в среднем 2% нейронов содержат NO-синтазу, в большинстве отделов головного мозга число таких нейронов также невелико. Однако имеются области и с высоким их содержанием. Так, максимальное количество нейронов, содержащих NO-синтазу, находится в коре мозжечка, где большинство клеток-зерен и корзинчатых нейронов содержат фермент [8]. Самые крупные нейроны коры мозжечка – клетки Пуркинье – не содержат NO-синтазу. Сравнительно много нейронов, содержащих NO-синтазу, находится в обонятельных луковицах, а также в некоторых отделах гиппокампа и полосатого тела. Только в мозжечке нейроны, содержащие NO-синтазу, составляют компактную популяцию клеток, а в остальных отделах – это одиночные, редко расположенные клетки (рис. 4) [2].

Рис.6. Нейроны, содержащие NO-синтазу, в коре больших полушарий белой крысы [2].
Обобщая имеющиеся данные о нейронах ЦНС, содержащих NO-синтазу, следует отметить, что преимущественная часть их относится к небольшим по размерам клеткам, многие из которых не содержат дендритных шипиков и являются ассоциативными нейронами. Крупные клетки, например пирамидные нейроны коры больших полушарий или гиппокампа, моторные нейроны передних рогов спинного мозга, не содержат NO-синтазу. NO-синтаза сосуществует в нервных клетках с другими традиционными нейромедиаторами и нейропептидами, чаще фермент определяется в холинэргических нейронах [2].
Большое внимание уделяется NO в реализации нервных воздействий на ткани внутренних органов. Нервы, содержащие NO-синтазу, показаны практически во всех изученных внутренних органах, преимущественно в стенке кровеносных сосудов, где они, наряду с эндотелием, могут оказывать сосудорасширяющий эффект [8] NO признается одним из основных эффекторных агентов в так называемых неадренэргических-нехолинэргических нервах. В периферических нервах подробно изучено сосуществование NO-синтазы с нейропептидами. Наиболее часто фермент определяется вместе с вазоактивным кишечнымнейропептидом (VIP) и нейропептидом Y [2].
Значение NO в ЦНС в нормальных условиях связывают с тремя процессами (так называемая NO-гипотеза):
1) Участие в межнейронной связи в качестве своеобразного нейромедиатора, причем основное значение, как полагают, NO имеет в синаптической пластичности, под которой понимают эффективность синаптической передачи;
2) Регуляция церебрального кровотока;
3) Установление межнейронных синаптических взаимосвязей во время развития нервной системы[2].
NO как нейромедиатор
Нейромедиаторная сущность NO заключается в том, что оно синтезируется при возбуждении нейрона (в ответ на поступление ионов кальция) и, диффундируя в соседние клетки, активизирует в них образование цГМФ, способного влиять на проводимость ионных каналов и, таким образом, изменять электрогенез нейронов [2]. NO отличается от традиционных нейромедиаторов тем, что он оказывает воздействие на ионные каналы не через плазмалеммальные рецепторы, а изнутри, со стороны цитоплазмы. Кроме того, действие NO не ограничивается только областью синаптических контактов, газ может влиять на ионные каналы на значительной площади плазматической мембраны нейрона [11].
Участие NO в синаптической пластичности наиболее ярко проявляется в таких процессах, как длительная синаптическая потенция (повышение эффективности проведения возбуждения через синапс для каждого последующего импульса в их последовательности), более детально проанализированная в гиппокампе, и длительная синаптическая депрессия (снижение эффективности проведения возбуждения через синапс), лучше исследованная в коре мозжечка[2].
В первом случае возбуждение постсинаптического окончания приводит к повышению внутриклеточного уровня Ca2+/кальмодулина, активирующих NO-синтазу и образование NO. NO, диффундируя в пресинаптическое окончание, вызывает образование цГМФ, что приводит к усилению и увеличению длительности выделения из пресинапса нейромедиатора, который оказывает возбуждающее действие на постсинаптический нейрон, и таким образом возникает обратная положительная связь с постоянным усилением. Такому механизму особое влияние уделяется в синапсах с наиболее распространенными и сильновозбуждающими нейромедиаторами ЦНС глутаматом и особому глутаматному рецептору – NMDA- рецептору, являющемуся трансмембранным каналом для ионов кальция (рис 7) [2].
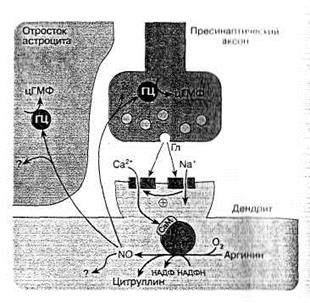
Рис. 7. Схема возбуждающего глутаматного синапса ЦНС и возможное участие NOв его деятельности [2].
Выделяющийся из синаптических пузырьков возбуждающий нейромедиатор глутамат (Гл) влияет на АМРА (А)-рецепторы, что приводит к деполяризации плазматической мембраны и открытию NMDA(N)-каналов (последнее связано с высвобождением вследствие изменения потенциала мембраны из ионного канала ионов магния, которые закрывали канал). Через NMDA-каналы в клетку поступает большой объем ионов кальция, активирующих конститутивную изоформу NO-синтазы. Образующийся NO, диффундируя в пресинаптический аксон, активирует в нем гуанилатциклазу (ГЦ) и синтез цГМФ, последний способствует усилению выделению нейромедиатора глутамата, что повышает эффективность синаптической передачи (феномен длительной синаптической потенции). NO может также влиять на окружающие астроциты и активировать в них гуанилатциклазу.
Дополнительные обозначения: СаМ – кальмодулин; ? – возможное влияние NO на другие не связанные с гуанилатциклазой клеточные системы нейронов и астроцитов.
С длительной синаптической потенцией, прежде всего в гиппокампе, связывают пластичность межнейронных связей, лежащих в основе памяти. Такое предположение основывается на ставших уже классическими представлениях Д.Хэбба (1949) о повышении эффективности синаптической передачи при возбужденном состоянии постсинаптического нейрона. Физиологические наблюдения об участии NO в процессах памяти и обучения противоречивы. Наряду с работами, в которых показано нарушение процессов обучения у экспериментальных животных при введении ингибиторов NO-синтазы, имеются и исследования с противоположными результатами. Неоднозначность получаемых результатов, возможно, связана с тем, что вводимые в организм ингибиторы NO-синтазы оказывают действие не только на всю нервную систему, но и на все органы и ткани животных, что не позволяло достичь локального влияния на продукцию NO в мозге [2].
В переживающих (живущих некоторое время в условиях культуры тканей) срезах головного мозга был показан и другой механизм действия NO на пресинаптические окончания. NO способен инициировать выделение нейромедиатора дофамина из нейронов не посредством экзоцитоза, а путем трансмембранной диффузии при участии особого мембранного переносчика дофамина, участвующего в его захвате из межклеточной среды [8]. В этом процессе не участвует цГМФ и предполагается прямое влияние NO на транспортные белки мембран [2].