Пептиды и первичная структура белка
Рис.19 Определение N-концевого остатка пептида. Пептид метят фтординитробензолом (реактив Сэнгера) и затем гидролизуют. ДНФ-производное аминокислоты (в приведенном примере ДНФ-аланин) идентифицируют по хроматографиче-ским характеристикам.
Для определения в белке или пептиде концевого остатка, несущего аминогруппу, его метят с помощью соединения, образующего стабильную ковалентную связь с азотом аминогруппы (рис. 20).
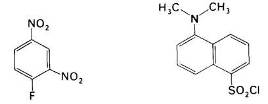
Рис. 20 фтординитробензол и дансилхлорид
Впервые для этой цели Сэнгер использовал фтординитробензол (ФДНБ), реагирующий с незаряженной a-NH2-группой с образованием динитрофенильного (ДНФ) производного пептида желтого цвета. Связь между ДНФ и концевой аминогруппой стабильна в условиях, используемых для гидролиза пептидных связей. Поэтому при гидролизе ДНФ-производного пептида Ala-Gly-Asp-Phe-Arg-Gly в 6 н. НСl высвобождается ДНФ-аминокислота, которую можно идентифицировать хроматографически как ДНФ-аланин.
Для идентификации N-концевых аминокислот в настоящее время часто используют дансилхлорид, который при взаимодействии с аминогруппой дает стабильное, интенсивно флуоресцирующее сульфамидное производное. Этот метод позволяет выявить N-концевую аминокислоту (после кислотного гидролиза пептидных
связей), присутствующую в таком незначительном количестве, как несколько нанограммов.
При всех достоинствах методов определения N-концевых аминокислотных остатков с помощью ДНФ или лансилхлорида их, к сожалению, нельзя использовать дважды применительно к одному и тому же пептиду, поскольку последний полностью распадается при кислотном гидролизе. Пьеру Эдману (P. Edman) удалось разработать метод маркирования N-концевого остатка и отщепления его от пептида без сопутствующего расщепления остальных пептидных связей. Деградация по Эдману (реакция Эдмана) состоит в ступенчатом (по одному) отщеплении аминокислотных остатков с аминоконца пептида, Фенилизотиоциа-нат реагирует с незаряженной концевой аминогруппой пептида с образованием фенилтиокарбамоильного производного
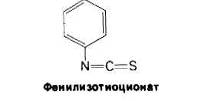
Далее в слабокислой среде происходит отщепление циклического производного N- концевой аминокислоты, а оставшийся неразрушенным пептид оказывается укороченным на один аминокислотный остаток, Указанное циклическое производное представляет собой фенилтиогидантоинаминокислоту (ФТГ-аминокислоту). Его идентифицируют методом хроматографии. Далее аминокислотный состав укороченного пептида (Arg, Asp, Gly2, Phe) сравнивают с исходным: (Ala, Arg, Asp, Gly2, Phe).
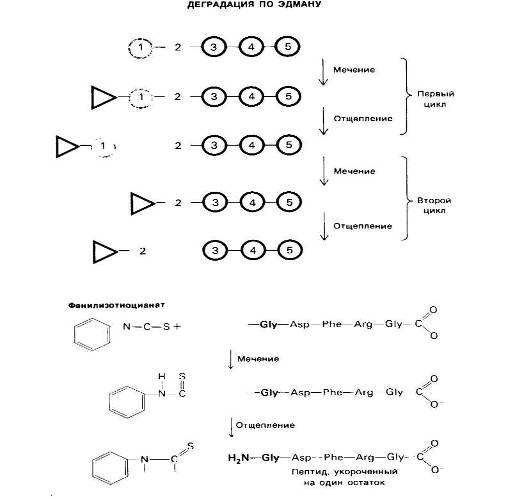 |
От пептидной цепи отщепляют меченый N-концевой остаток аминокислоты (ФТГ-аланин на первой ступени деградации). Остаток пептидной цепи при этом не гидролизуется. На второй ступени деградации определяют следующий N-концевой аминокислотный остаток. Еще три ступени деградации но Эдману позволят установить всю последовательность аминокислот во взятом пептиде.
Оказывается, что различие состоит в одном остатке аланина. Следовательно, в исходном пептиде аланин занимает N-концевое положение. Деградацию по Эдману можно вновь повторить па укороченном пептиде. Исходя из аминокислотного состава после второй ступени деградации
(Arg, Asp, Gly, Phe)
Можно поийти к выводу, что вторым остатком с N-конца является глицин, это заключение подтверждают путем хроматографической идентификации ФТГ-глицина, полученного на второй ступени деградации пептида. Еще три ступени деградации по Эдману позволяют полностью раскрыть последовательность аминокислот во взятом пептиде.
Стратегию анализа последовательности аминокислот в белках можно определить как «разделяй и властвуй». Белок подвергают специфическому расщеплению па более короткие пептиды, последовательность аминокислот в которых определяют по Эдману. Специфическое расщепление можно производить химическими или ферментативным методами. Так, Б. Уиткоп (В. Witkop) и Э. Гросс (Е. Gross) обнаружили, что бромистый циан (CNBr) расщепляет полипептидную цепь только по пептидной связи, образованной карбоксильной группой остатка метионина, Если в белке содержится 10 метиониновых остатков, то после обработки бромистым цианом обычно получается 11 пептидов. Высокоспецифическое расщепление достигается также с помощью трипсина-протеолитического фермента поджелудочной железы.
Трипсин расщепляет полипептидные цепи по пептидной связи, образованной карбоксильной группой остатков аргинина и лизина. В результате белок, содержащий 9 остатков лизина и 7 остатков аргинина, после расщепления трипсином распадается на 17 пептидов. Каждый из этих пептидов, кроме пептида, расположенною на карбоксильном конце белка, будет кончаться аргинином или лизином.
Пептиды, полученные при специфическом химическом или ферментативном расщеплении белка, разделяют методами хроматографии. Далее последовательность аминокислот в каждом из пептидов определяют методом Эдмана. Таким образом, достигается этап, когда последовательность аминокислот в отдельных пептидах (фрагментах белка) известна, но остается невыясненной последовательность самих пептидов. Последнюю устанавливают с помощью так называемых перекрывающихся пептидов. При этом используют уже не трипсин, а какой-либо фермент, расщепляющий полипептидную цепь в других участках, например химотрипсин, который расщепляет пептидные связи главным образом по карбоксильным группам ароматических и других больших неполярных аминокислотных остатков
|
|
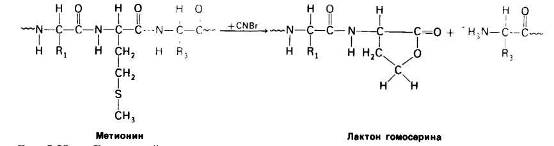
Рис. 23 Бромистый циан расщепляет полипептиды по карбоксильной группе метиониновых остатков.
Пептиды, образующиеся под действием химотрипсина, неизбежно перекрывают два или более триптических пептида, что используется для установления их последовательности. Таким путем полностью определяют последовательность аминокислот в белке.
Описанные методы применимы к белкам, состоящим из одной полипептидной цепи, не имеющей дисульфидных связей. В тех же случаях, когда в белке имеются дисульфидные связи или более одной полипептидной цепи, то необходимы дополнительные методические приемы, Например, если белок содержит две или более полипептидные цепи, соединенные нековалентными связями, то, воздействуя денатурирующими агентами, такими, как мочевина или гуанидингидрохлорид, вызывают диссоциацию цепей. Диссоциированные цепи разделяют и только после этого приступают к определению последовательности аминокислот в каждой из них. Если же полипептидные цепи соединены ковалентными дисульфидными связями, как это имеет место в инсулине, то их окисляют надмуравьиной кислотой; при этом дисульфидные связи разрываются и образуются остатки цистеиновой кислоты.