Строение, свойства и биологическая роль биотина и тиамина
Серия работ по изучению тиаминкиназы, выделенной из дрожжей и животных тканей, недавно проведена в Японии. На достаточно очищенных препаратах (более чем в 100 раз), обладавших слабой АТФ-азной активностью и не содержащих аденилаткиназы и нуклеозиддифосфокиназы, установлено, что ионы марганца, магния - кобальта активировали, а кальция, никеля, рубидия и железа — в широком диапазоне концентраций не угнетали фермент. На этом же препарате фермента показаны возможность фосфорилирования тиамина за счет других нуклеотидтрифосфатов (ГТФ, ИТФ, УТФ и др.) но то, что основным продуктом реакции является ТДФ и небольшое количество ТМФ. Применением Р32-АТФ, как и в исследованиях предыдущих авторов, подтвержден механизм переноса на тиамин сразу пирофосфатной группировки.
Однако результаты, полученные in vitro, не нашли полного подтверждения при изучении фосфорилирования тиамина на целых организмах и в опытах с митохондриями. С одной стороны, как будто подтверждалось предположение о пирофосфорилировании: после внутривенного введения тиамина уже через 30—60 минут в крови животных обнаруживались меченные по фосфору ТДФ и ТТФ, но не ТМФ. С другой стороны, после внутривенного введения ТМФ кокарбоксилазная и транскетолазная (Н. К. Лукашик, 1964) активность крови нарастала быстрее, чем после введения свободного тиамина. Некоторые микроорганизмы легче образуют ТДФ из ТМФ, чем из свободного витамина, а тиаминкиназа, найденная ранее в печени, не обнаружена в митохонд-риях почек, в которых фосфорилирование тиамина идет другим путем. Механизм фосфорилирования витамина с участием только АТФ не всегда укладывается в простую схему переноса пирофосфатной группировки в целом хотя бы потому, что наряду с ТДФ в различном биологическом материале обнаруживаются в значительных количествах и другие ТФ, в том числе даже Т-полифосфаты.
Ряд исследований последних лет касается вопроса о локализаций систем, ответственных за фосфорилирование тиамина. Печень уже через час после введения тиамина захватывает 33—40% витамина, накапливая различные его фосфорные эфиры. По данным А. А. Рыбиной (1959), происходит быстрое фосфорилирование меченого витамина и в других органах (в порядке убывающей активности): печень, почки, сердце, семенники, головной мозг. При этом радиоактивность фосфорных эфиров тиамина убывает в ряду: ТТФ, ТДФ, ТМФ. По данным японских авторов, фосфорилирование тиамина идет активно в митохондриях (Shima-zono, 1965), микросомах и гиалоплазме.
Из изложенных выше фактов нетрудно сделать вывод, что общая -интенсивность процессов эстерификции витамина в организме или в отдельных тканях должна в значительной степени коррелировать с активностью процессов, поставляющих АТФ. Первые экспериментальные наблюдения в этом плане, проведенные ня гомогенатах печени или клеточных элементах крови, получили в дальнейшем полное подтверждение. Все яды дыхания и гликолиза или соединения, конкурирующие с Т за АТФ, как правило, снижают уровень ТДФ в крови и в тканях.
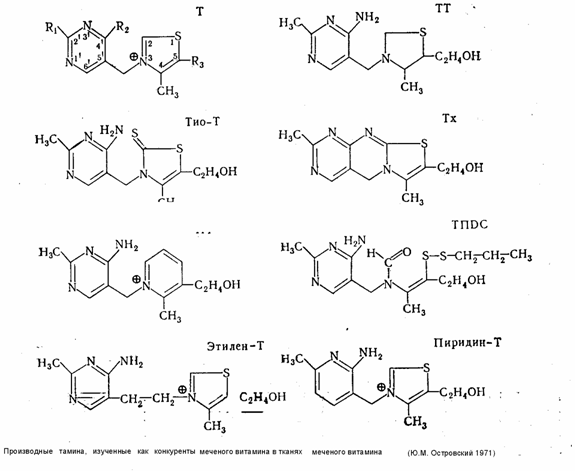
4.2 Роль отдельных группировок в молекуле тиамина для его связывания в тканях.
За последние годы синтезировано более десятка новых производных тиамина (смешанные дисульфиды, О-бензольные производные и др.), широко внедряемых в лечебную и профилактическую практику. Преимущества новых витаминных препаратов, как правило, выявлялись чисто эмпирически в связи с тем, что до настоящего времени мы не располагаем достаточными сведениями о молекулярных механизмах ассимиляции тиамина, о характере его взаимодействия со специфическими (ферменты) и неспецифическими (осуществляющими транспорт витамина) белками. Необходимость точных представлений в этом вопросе диктуется и широкими перспективами использования антивитаминов тиамина (ампрол, хлоротиамин, деокситиамин) для лечебных целей (см. ниже).
Работы по синтезу новых производных тиамина с заранее заданными физико-химическими свойствами, обусловливающими возможности целенаправленного воздействия на обменные процессы в организме, немыслимы без конкретных представлений о роли отдельных групп атомов витамина и его производных в этой области. Значение пирофосфатного радикала для специфической потеидизации ТДФ в составе соответствующих ферментов уже отмечалось выше. В последние годы становится все более очевидным участие тиамина в других реакциях, не имеющих ничего общего с коферменными функциями витамина. Разнообразию активных группировок в молекуле тиамина соответствует каждый раз особая форма- претеидизации, блокирующая одни и обнажающая одновременно другие, важные для соответствующей функции, участки молекулы витамина (Ю. М. Островский, 1971). Действительно, первый тип протеидизации (через пирофосфатный радикал) отвечает коферментной функции и оставляет свободными, доступными для субстрата 2-й углерод тиазола и аминогруппу пиримидинового компонента. С другой стороны, очевидно, что участие витамина в окислительно-восстановительных реакциях или в процессах перефосфорилирования должно сочетаться с исключением возможности одновременного функционирования его как кофермента, так как в первом случае необходима деполяризация и раскрытие тиазолового цикла, а во втором — свободное положение фосфорилированного оксиэтильного радикала. Поскольку 80—90% тиамина, присутствующего в тканях, освобождается лишь при кислотном и ферментативном гидролизе, можно считать, что все связанные формы витамина находятся в протеидизированном, т. е. связанном с белками, состоянии.
Представление о значении отдельных участков молекулы тиамина в этом процессе легко получить, определяя степень связывания тканями меченного по сере (S35) витамина и некоторых его производных, лишенных тех или иных активных центров, например аминогруппы — окситиамин (окси-Т), аминогруппы и оксиэтильного радикала — хлорокситиамин (ХОТ), четвертичного азота в тиазоловом цикле тетрагидротиамин (TТ) Некоторые ограничения на интерпретацию данных, полученных таким образом в короткие сроки опыта, накладывают физиологические механизмы, транспорта и экскреции вводимых соединений, в связи с чем дополнительно приходится исследовать также выведение витамина и самих меченых соединений с мочой. С другой стороны, результаты, получаемые в длительные сроки (24 часа), почти полностью соответствуют только представлениям о протеидизированной части витамина, а на основании рассмотрения конкурентных взаимоотношений между различными упомянутыми выше мечеными и другими немечеными производными витамина можно последовательно исключать роль отдельных атомов или группировок в механизмах фиксации тиамина тканями.
4.3 Коферментные функции тиаминдифосфата.
Известно значительное количество различных реакций, катализируемых ТДФ. Однако все их можно свести к нескольким типичным вариантам: простое и окислительное декарбоксидирование a-кетокислот, ацилоиновая конденсация, фосфорокластическое расщепление кетосахаров. Ферментные системы, принимающие участие в этих реакциях по-видимому, едины в основных принципах своего действия; различна лишь последующая судьба «активного альдегидного осколка», возникающего на первых этапах процесса. Успехи, достигнутые в течение последних лет в изучении превращений a-кетокислот (С. Е. Северин, 1964; А. А. Глемжа, 1964), позволили четко представить как роль собственно декарбоксилирующего фрагмента полиферментного комплекса дегидрогеназы, содержащего ТДФ, так и последовательность всех других, связанных с ним реакций: