Мембранная энзимология
Присутствие фосфолипидов сильно меняет стационарную кинетику реакций протеолитической активации. Резко уменьшается значение Км для белкового субстрата; например, для реакции активации фактора X фактором 1Ха Км изменяется от 181 до 0,058 мкМ. Добавление другого белка, фактора Villa, увеличивает Kma* более чем в 200 000 раз. Поскольку реакция катализируется обоими ферментами, а субстрат в данных условиях измерения представлен как мембраносвязанной, так и свободной формами, истинный механизм влияния липида в таких реакциях определить чрезвычайно сложно. Например, показано, что протеолитическая активность фактора X увеличивается при связывании его с мембраной, в то время как фактор IX одинаково активен в свободном и в мембраносвязанном состояниях. Фактор X можно также активировать комплексом фактора Vila и тканевого фактора. В этом случае протеолитическая активация фактора X происходит, только когда он находится в свободной форме в растворе и не связан с мембраной. Другой пример - активация протромбина фактором Ха. Наблюдаемое в этом случае низкое значение Км коррелирует с концентрацией субстрата и протромбина на поверхности фосфолипидной везикулы. Если же добавить кофактор - фактор Va, образующий с фактором Ха комплекс, - то всякая зависимость Км от поверхностной концентрации протромбина на везикуле исчезнет. В заключение отметим, что данная система очень сложна, и роль липидов здесь отнюдь не сводится лишь к созданию соответствующей поверхности, на которой происходит простое концентрирование компонентов системы.
Факторы свертывания крови входят в группу Са2 +-зависимых липидсвязывающих белков. Функции этих белков не всегда бывают известны; некоторые из них связаны с цитоскелетом. Фосфолипазы, к рассмотрению которых мы сейчас перейдем, также являются Са2 +-зависимыми ферментами.
6.6 Фосфолипазы - растворимые ферменты, катализирующие расщепление мембраносвязанных субстратов
Фосфолипиды служат субстратами многих растворимых ферментов, в том числе фосфолипаз. Среди них лучше всего изучена фосфолипаза Аг, которая катализирует гидролиз фосфолипидов по положению sn-2 с образованием жирной кислоты и лизофосфолипида. Фосфолипаза Аг была выделена сначала из ядов кобры и гремучей змеи, а затем из поджелудочной железы быка и свиньи. Это очень близкие по первичной структуре небольшие белки с мол. массой около 14 000. Для некоторых ферментов удалось получить с высоким разрешением трехмерные структуры, также обладающие высокой степенью гомологии. Ферменты из поджелудочной железы синтезируются как неактивные зимогены, которые затем активируются протеолизом: от зимогена отщепляется семь остатков с С-конца.
Фосфолипаза Аг представляет особый интерес с точки зрения мембранной энзимологии, поскольку она обладает способностью активироваться при взаимодействии с интегрированными формами субстрата, например с мицеллами или бислоем. На Рис.6.8 представлена зависимость от концентрации субстрата скорости гидролиза короткоцепочечного фосфатидилхолина фосфолипазой Аг и его предшественником из поджелудочной железы свиньи.
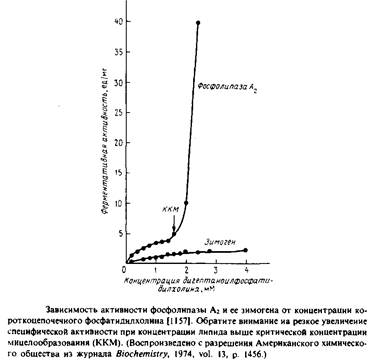
Данный субстрат в концентрациях до 1,5 мМ является мономером, но при дальнейшем увеличении концентрации формирует мицеллы. И зимоген, и активированный фермент очень медленно гидролизуют субстрат в мономерной форме, но как только фосфолипид начинает образовывать мицеллы, активность фосфолипазы А2 резко возрастает.
Активации фосфолипазы агрегированными субстратами было посвящено множество работ, а которых исследовалась кинетика катализируемого ферментом гидролиза субстратов в мономерной форме, в чистых липидных мицеллах, в смешанных мицеллах с тритоном Х-100, в монослоях на поверхности раздела воздух-вода и в фосфолипидных везикулах. Для проявления каталитической активности ферменту во всех случаях нужен Са2 +, причем центр связывания единственного иона Са2+ можно выявить с помощью рентгеноструктурного анализа. В отличие от факторов свертывания крови фосфолипаза Аг не содержит остатка - у-карбоксиглутаминовой кислоты и для ее активации не требуются кислые фосфолипиды.
Для объяснения механизма активации фосфолипаз предложено несколько гипотез
В ряде работ было показано, что связывание фермента с мицеллами или бислоями предшествует стадии активации, при которой резко возрастает число оборотов фермента, и экспериментально эти две стадии можно разделить. Такое поведение ничем не отличается от поведения других рассмотренных липидзависимых ферментов. Несмотря на обилие данных по кинетике, связыванию и структуре фосфолипазы, исследователи не пришли к единому мнению о том, что происходит с ферментом при его активации в присутствии липидного бислоя или мицелл. В литературе рассматривается несколько возможных механизмов.
Фермент связывается с бислоем с помощью специального "участка узнавания поверхности раздела", отличного от активного центра, и для его формирования необходим Са2 +. Предполагается, что этот участок проникает в глубь мембраны. Эта модель основана, в частности, на данных по специфическому влиянию химической модификации N-концевого участка полипептида на взаимодействие с агрегированными субстратами. Происходящая при взаимодействии участка узнавания с мембраной активация фермента, по-видимому, обусловлена конформационными изменениями белка. Следует отметить, что в кристаллическом виде ферменты из поджелудочной железы быка и свиньи представляют собой мономеры, в то время как фосфолипаза Аг из яда гремучей змеи является димером. Обнаруживаемый в мономерных фосфолипазах участок, который, как предполагают, является "участком узнавания поверхности", в димерном ферменте недоступен из водной фазы и находится на поверхности межсубъединичного контакта.
Двухфосфолипидная модель предполагает существование в ферменте двух или более центров связывания фосфолипидов и основана прежде всего на кинетических данных по активации фермента фосфолипидами в смешанных мицеллах. Эта модель позволяет учесть роль агрегации двух или более молекул фермента как важнейшей части схемы активации, а также роль возможных конформационных изменений в увеличении каталитической активности.
Постулируется, что конформация фосфолипидного субстрата в агрегированном состоянии отличается от конформации мономерной формы, и именно с этим связана более высокая скорость гидролиза агрегированных форм липидов ферментом.
4. Увеличение активности связано с тем, что из мицелл или бислоя продукты гидролиза удаляются легче. Кроме того, само по себе накопление продуктов уже приводит к увеличению активности фосфолипазы Аг, хотя механизм этого явления неясен.
Одна из проблем, возникающих при анализе процесса активации, состоит в том, как разделить процессы связывания с липидом и активацию липидом. В экспериментах с однослойными фосфолипидными везикулами удалось выяснить, что критическим параметром для обоих стадий является физическое состояние бислоя. Показано, например, что фосфолипаза А2 лучше всего связывается с дипальмитоилфосфатидилхолином в фазе геля, причем Са2 + для этого не нужен. Для активации же фермента в такой системе, по-видимому, требуется Са2 +, причем в случае везикул фосфатидилхолиновый бислой должен обладать дефектами упаковки и в нем должны происходить структурные флуктуации, подобные тем, которые имеют место в ходе термоиндуцируемого фазового перехода. Взаимодействия молекул белков могут быть важны как для связывания, так и для активации. В некоторых условиях активированный фермент сохраняет активность по крайней мере в течение 30 мин.