Мембранная энзимология
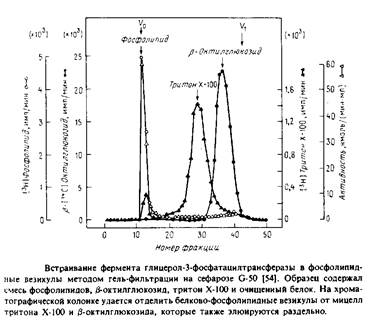
С помощью такой методики можно ввести в везикулы отличные от фосфолипидов вещества, например холестерол или убихинон. В ряде случаев возникает необходимость в достаточно быстром удалении детергента, особенно если белок нестабилен при его избытке. Тогда применяют гельфильтрационную хроматографию, тоже позволяющую эффективно отделить белково-липидные везикулы от детергента. На рис.1 показан профиль элюции с такой колонки.
Еще более быстрым является метод разведения, когда белково-липидно-детергентную смесь разводят до концентрации детергента много меньшей, чем критическая концентрация мицеллообразования. При этом спонтанно образуются белково-фосфолипидные везикулы, которые можно отделить от детергента центрифугированием.
Обычно используемый для очистки мембранных ферментов тритон Х-100 весьма неудобен при реконструкции, поскольку его трудно Удалить из системы. Для удаления тритона применяют шарики из полистирола, и одна из проблем - потеря белка из-за его сорбции на поверхности шариков. В качестве примера белка, реконструируемого этим методом, можно привести натриевый канал.
И наконец, для реконструкции применяли метод, основанный на диспергировании смеси холестерол-фосфолипид-белок в диэтиловом эфире и последующем испарении обращенной фазы.
Однако маловероятно, что многие белки выдержат такую процедуру.
3. Некоторые характеристики реконструированных белково-фосфолипидных везикул
В результате применения всех методов, разработанных для реконструкции мембранных белков, образуются однослойные везикулы. Характеристики некоторых из них детально изучены. Это, в частности, протеолипосомы, содержащие цитохром с-оксидазу, Ыа + /К+-АТРазу, цитохром Ь5 и гликофорин. Особый интерес представляют:
1) средний размер везикул и распределение их по размерам;
2) распределение белка в популяции везикул;
3) ориентация белка по отношению к плоскости бислоя;
4) проницаемость везикул. Эти характеристики особенно важны, если фермент катализирует трансмембранный перенос веществ или ионов. Для количественного анализа кинетики таких ферментов необходимо знать внутренний объем везикул, их проницаемость и распределение белка. Для ферментов, катализирующих векторные реакции, молекулы, которые имеют противоположную ориентацию и находятся в одной везикуле, будут работать "вхолостую", компенсируя друг друга при создании результирующего ионного градиента.
Размер везикул. Размер везикул сильно зависит от процедуры реконструкции. Для его определения лучше всего использовать методы электронной микроскопии или гельфильтрации. При удалении детергента диализом образуются протеолипосомы диаметром от 500 до 2500 А в зависимости от белка и используемого метода. При любом способе реконструкции размеры получаемых везикул варьируют в довольно широких пределах; протеолипосомы нужного диаметра можно затем отделить с помощью гельфильтрационной хроматографии.
Распределение белка. Когда при реконструкции используется диализ для удаления детергента из смеси, содержащей избыток ли-пида, распределение белка между полученными везикулами должно подчиняться распределению Пуассона. Однако на самом деле оно может зависеть от встраиваемого белка и методических деталей. Следует отметить, что при спонтанном встраивании в предварительно полученные липосомы некоторые белки включаются преимущественно в везикулы малых размеров. Цитохром bs, например, в 200 раз эффективнее встраивается в везикулы диаметром 200 А, чем 1000 А.
Ориентация белка. Этот вопрос важен с точки зрения энзимологии, поскольку белок, активный центр которого локализован внутри везикулы, может быть недоступен для субстрата. Как ни удивительно, многие ферменты, например цитохром с-оксидаза, Ыа + /К+-АТРаза, глицерол-3-фосфатацил-трансфераза, бактериородопсин, способны встраиваться в мембрану таким образом, что их активный центр с вероятностью 75-95Чо оказывается снаружи везикулы. Везикулы, содержащие цитохром с-оксидазу, с помощью ДЭАЭ-хроматографии удается разделить на две популяции: с активным центром, ориентированным внутрь, и с активным центром, ориентированным наружу. В принципе такой способ разделения пригоден и для других белков.
Причина такой асимметрии встраивания неизвестна. В некоторых случаях ориентированная наружу часть фермента имеет больший размер, а встраивание белка более массивной частью внутрь везикулы невыгодно. Поскольку преимущественная ориентация наблюдается и при встраивании белков в крупные липосомы, по-видимому, асимметрия встраивания не связана с кривизной везикулы, как в случае распределения липидов в везикулах малых размеров. Вероятно, важную роль играют какие-то кинетические факторы, однако их трудно оценить, поскольку неясна природа переходных состояний.
Нельзя не отметить также, что некоторые белки встраиваются в везикулы неправильным образом, т.е. в конформации, отличной от нативной. Наиболее типичный пример - цитохром bs, который в зависимости от метода реконструкции может находиться в одной из двух конформации. Такое поведение характерно также для компонента Н-2К главного комплекса гистосовместимости у мышей и, возможно, для белка оболочки бактериофага Ml 3. Все эти белки имеют один трансмембранный гидрофобный домен, и при некоторых условиях он включается в бислой в U-образной конфигурации, когда N - и С-концевые аминогруппы экспонированы наружу.
Проницаемость. Важность этой характеристики для ферментов, катализирующих перенос веществ или ионов через бислой, несомненна. Многие системы транспорта и ионные насосы изучали после встраивания их в протеолипосомы, и очевидно, что протеолипосомы, пригодные для таких исследований, должны обладать достаточно низкой проницаемостью. Присутствие белка обычно приводит к увеличению проницаемости везикул, но степень этого увеличения очень сильно зависит от выбора липида и от числа молекул белка на везикулу. Высказывалось предположение, что молекулы некоторых липидов благодаря их форме лучше упаковываются вокруг встроенных в бислой белков; тем самым сглаживаются дефекты структуры на границе белок-липид и уменьшается их проницаемость для растворенных веществ. Однако прямые доказательства по этому поводу отсутствуют.
3.1 Влияние липидов на активности мембраносвязанных ферментов
Каталитическая активность многих мембранных ферментов зависит от липидов. Липиды при этом могут выполнять две функции:
1) создавать необходимую среду;
2) действовать как аллостерический регулятор, модулирующий активность фермента. В первом случае липиды не только предотвращают денатурацию ферментов, но и облегчают взаимодействие ферментов друг с другом и с прочими мембраносвязанными компонентами, в частности с ли-пофильными субстратами. В качестве аллостерических эффекторов липиды обычно активируют фермент преимущественно путем стабилизации его в определенной конформации. В принципе две эти функции липидов совершенно различны, однако разграничить их экспериментально бывает крайне трудно. В идеальной экспериментальной системе аллостерическим эффектором должен быть специфический липид, а необходимое для работы окружение фермента должно создаваться всей основной массой липидов в бислое. К сожалению, пока обнаружен только один пример абсолютной специфичности фермента к определенному липиду. /З-Гидроксибутиратде-гидрогеназе для проявления каталитической активности необходим именно фосфатидилхолин, в большинстве же случаев ферменты достаточно эффективно активируются различными липидами. А поскольку любой липид может в той или иной степени выполнять как функцию окружения, так и функцию специфического аллостерического эффектора, различить эти два эффекта становится практически невозможно. На ферментативную активность может влиять также физическое состояние бислоя, в частности поверхностная плотность заряда и вязкость. Систематически подбирая липиды разной структуры и изменяя физическое состояние мембраны, можно установить корреляции между активностью фермента и этим" параметрами, но само по себе такое исследование еще не позволяет разграничить две функции липидов. Изменение физического состояния бислоя может влиять на взаимодействие с ферментом любых липидов, в том числе и тех, которые функционируют как аллостерические эффекторы. При изучении ферментов, субстратом которых служат липофильные соединения, возникают особые проблемы, поскольку такие субстраты должны быть включены в бислои до или после получения везикул, а большие концентрации растворенного в бислое субстрата не могут не сказаться на физическом состоянии модельной мембраны.