Мембранная энзимология
На Рис.6.2 приведена кривая активации БДГ фосфолипидными везикулами, содержащими разное количество фосфатидилхолина. Во всех случаях фермент встроен в липосомы. Зависимость скорости ферментативной реакции от содержания фосфатидилхолина в бислое указывает на высокую кооперативность процесса, в котором участвуют два необходимых для активации липидсвязывающих центра.
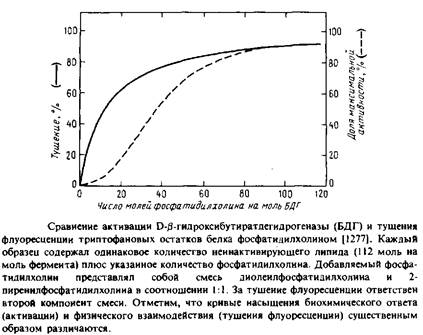
Кроме того, по данным о тушении флуоресценции триптофана производными фосфатидилхолина отдельно была определена доступность молекул фосфатидилхолина для БДГ. Степень тушения флуоресценции фактически является мерой связывания липида. Оказалось, что фосфатидилхолин связывается некооперативно с примерно 12 центрами в молекуле фермента. Такое некооперативное связывание с довольно большим числом центров весьма типично для мембранных белков в бислое. Кроме того, основная часть флуоресценции тушится при более низких концентрациях фосфатидилхолина, чем необходимо для активации. Такое расхождение между связыванием липида и активацией фермента можно объяснить, если предположить, что в активации участвует только очень небольшая часть центров, которые не удается выявить в экспериментах по тушению флуоресценции. Приведенные результаты интересны еще и тем, что позволяют сопоставить физическое взаимодействие определенного липида и фермента с биохимическим ответом.
Интерпретация результатов таких экспериментов осложняется тем, что активность фермента может сильно зависеть от степени его агрегации, т.е. активность фермента в различных липидах, по-видимому, будет соответствовать степени его дезинтеграции. БДГ замечателен еще и тем, что это один из немногих ферментов, активируемых короткоцепочечными гомологами фосфатидилхолина, которые связываются с ферментом и активируют его в концентрациях ниже критической концентрации мицеллообразования. Эти данные тоже подтверждают, что в активацию липидом вовлечено только небольшое число центров связывания на белке.
4.2 Пируватоксидаза
Эта флавиносодержащая дегидрогеназа из Е. coli, катализирующая окисление пирувата до уксусной кислоты и СОг и восстановление убихинона в цитоплазматической мембране, обеспечивает поступление электронов в аэробную дыхательную цепь. Фермент обладает рядом замечательных особенностей. Он растворим в воде и не проявляет никаких характерных для мембранного белка свойств. Однако в присутствии субстрата и кофактора происходит изменение конформации белка, в результате чего формируется центр связывания с мембраной. В этих условиях резко возрастает сродство фермента к детергентам и фосфолипидным везикулам, и белок по своим свойствам становится похож на истинный мембранный фермент. Пируватоксидаза может служить примером фермента, который является цитоплазматическим, пока субстрата мало, но превращается в мембранный, как только концентрация субстрата становится достаточно высокой.
Наибольший интерес при исследовании этого фермента представляла его эффективная активация липидами. Активность фермента можно измерить, используя вместо природного мембраносвязанного акцептора - убихинона какой-либо растворимый в воде искусственный акцептор электронов, например феррицианид. В присутствии липидов каталитическая активность пируватоксидазы возрастает до 50 раз. В отличие от /3-гидроксибутиратдегидрогеназы этот фермент активируется самыми разнообразными фосфолипидами и детергентами. В концентрациях ниже критических концентраций мицеллообразования фермент полностью активируется как анионными, так и катионными детергентами, а исследование связывания детергентов показывает, что в активации участвует очень небольшое число центров связывания. Связывание фермента с фосфатидилхолиновыми везикулами или детергентами не происходит до тех пор, пока не добавлены субстрат и кофактор. В присутствии субстрата и кофактора пируватоксидаза способна связываться и активироваться разнообразными везикулами, полученными из целого ряда фосфолипидов. Связывание происходит в первую очередь за счет гидрофобных, а не электростатических взаимодействий. Связывание липида и активация в случае пируватоксидазы, по-видимому, неразделимы, хотя и не все везикулы связываемого фосфолипида или амфифильного соединения ответственны за активацию фермента. Активация липидами вызывает изменения в спектре поглощения связанного флавина, вероятно, вследствие облегчения реакции переноса электронов.
Конформационные изменения в молекуле пируватоксидазы приводят не только к образованию липидсвязывающего домена, но и к появлению чувствительного к протеазам участка цепи вблизи С-конца. Протеолиз подавляет связывание липидов, но, как это ни удивительно, вызывает активацию фермента в отношении реакции восстановления водорастворимых искусственных акцепторов электронов. Активированный протеазами фермент, однако, уже не может восстанавливать мембраносвязанный убихинон, поскольку утрачивает способность связываться с мембраной.
Как показывают результаты клонирования и секвенирования гена, кодирующего пируватоксидазу, аминокислотная последовательность белка не имеет протяженных гидрофобных участков. Генетические исследования, а также данные по протеолизу показали, что за связывание липида ответствен участок полипептида, локализованный вблизи С-конца. В этом участке имеется короткая потенциально амфифильная а-спираль, которая и может участвовать в связывании с поверхностью бислоя. Получен мутантный штамм Е. coli, у которого пируватоксидаза лишена последних 24 аминокислотных остатков. Такой фермент полностью неактивен in vivo, но in vitro успешно катализирует реакцию с водорастворимым акцептором электронов. Предположительно такой мутантный вариант пируватоксидазы не способен связываться с мембраной даже при высоких концентрациях субстратов, а значит, не может осуществлять катализ, поскольку для этого фермент должен окисляться убихиноном. Приведенное исследование является хорошим примером того, как данные по связыванию липида и активации, полученные in vitro, могут с успехом применяться для объяснения механизма функционирования фермента в клетке.
И пируватоксидаза, и /3-гидроксибутиратдегидрогеназа могут активироваться либо 1) при связывании с небольшим числом молекул амфифильного соединения, либо 2) при связывании с поверхностью бислоя. В последнем случае некоторая часть белка проникает в глубь мембраны. Оба рассмотренных примера четко иллюстрируют роль фосфолипидов как аллостерических регуляторов.
4.3 Са2 + - АТРАза
Если предыдущие два примера иллюстрируют роль липидов как аллостерических эффекторов мембранных белков, то Са2 + - АТРаза и еще два фермента, рассмотренные ниже, являются примерами ферментов, на активность которых влияют структура липида и физическое состояние бислоя. Выделенная из саркоплазматического ретикула скелетных мышц Са2+ - АТРаза состоит из единственной полипептидной цепи с мол. массой 115 кДа. Фермент осуществляет активный транспорт Са2+ внутрь саркоплазматического ретикулума, уменьшая тем самым концентрацию Са2 + в цитоплазме во время релаксации мышцы. На каждую гидролизованную молекулу АТР через мембрану переносятся два иона Са2 +, и реакция является электрогенной, т.е. перенос зарядов через бислой создает трансмембранный электрический потенциал. По результатам секвенирования соответствующего гена установлена аминокислотная последовательность белка. Предполагается, что полипептид имеет 10 трансмембранных спиральных участков.