НейрокомпьютерыРефераты >> Информатика >> Нейрокомпьютеры
В настоящее время установлено, что мозг, судя по всему, основан на принципе относительно жестко запаянного блока, состоящего из сложно организованных нейронных сетей, работающих в миллисекундном диапазоне. Более детальное изучение этих сетей осложняется специфическими свойствами нервной ткани, содержащей помимо нервных клеток и другие клетки, которые поддерживают нейрон механически и участвуют в процессах их метаболизма и проведения спайков.
В целом, нервная ткань представляет собой бесцветную студенистую массу, в которой даже под микроскопом трудно различить отдельные нейроны и состоящие из них сети. Поэтому в современной нейроанатомии применяют специальные методы окрашивания нервной ткани. В частности, используются красители, которые избирательно воздействуют лишь на некоторые нейроны и окрашивают их целиком. Окрашенная таким образом ткань замораживается, разрезается на тонкие слои и изучается под микроскопом. В процессе изучения удается выделить отдельные нейроны в сетях плотно упакованных нервных клеток, волокна которых тесно переплетены в густую чащу с промежутками 0,01 мкм. Более того, удается не только различать отдельные клетки, но и находить их связи друг с другом, как в локальных областях нервной ткани, так и в различных, далеко отстоящих друг от друга частях мозга. Однако получаемые при этом сведения не являются полными и не позволяют делать однозначных выводов о конфигурациях и законах функционирования изучаемых нейронных сетей. Эти сведения приходится дополнять данными других исследований, а именно тех, которыми занимается нейрофизиология.
Основным нейрофизиологическим подходом к исследованию мозга в настоящее время служит микроэлектродная методика. Ее суть заключается в том, что в живую ткань мозга вживляются микроэлектроды, с помощью которых регистрируется электрическая активность отдельных клеток. Однако использование этой методики для изучения высокоразвитых животных связано с рядом трудностей. Размеры электродов по сравнению с микроскопически малыми телами клеток велики, а возможности их миниатюризации ограничены. Поэтому микроэлектродные исследования могут искажать нормальную работу изучаемых структур. Вживление электродов осуществляется вслепую и не всегда ясно, работу каких клеток характеризуют снятые с них сигналы. Кроме того, клеток много, а электродов мало. Поэтому получаемая с их помощью информация является фрагментарной.
С целью преодоления отмеченных трудностей в нейробиологии вообще и в нейрофизиологии в частности широко применяют метод биологического моделирования. В соответствии с этим методом для изучения сложных структур мозга высокоразвитых животных используют достаточно простые нервные системы таких беспозвоночных, как черви, моллюски, кальмары, раки и т. п. Нейронные структуры беспозвоночных содержат сравнительно мало нейронов, упакованных в нервные узлы - ганглии. Ганглии, в свою очередь, содержат от 100 до 1000 клеток. Сами клетки у беспозвоночных крупнее, чем у млекопитающих, а их положение в ганглиях и межнейронные связи более определены.
Рассмотренный подход к изучению нервной системы внешне напоминает хорошо известный в технике метод физического моделирования. В соответствии с этим методом сложный для изучения объект заменяется менее сложным, но эквивалентным исходному по сути изучаемых явлений.
Однако в случае нейробиологии о собственно физическом моделировании можно говорить лишь в том случае, когда исследуются такие свойства нервной ткани, как электропроводность внутриклеточной среды, электрическая емкость и сопротивление мембран, механизм генерации спайков и т. п. Эти свойства, судя по всему, являются фундаментальными и не зависят от вида животного. В то же время механический перенос данных о структуре и свойствах нейронных сетей простых животных, например беспозвоночных, на нервную систему высокоразвитых млекопитающих вряд ли является корректным. Поэтому для изучения информационных процессов в сложных нейронных сетях необходимы современные методы математического и кибернетического моделирования. Причем результаты экспериментов на простых организмах могут использоваться в данном случае в качестве исходных гипотез для построения адекватных кибернетических моделей мозга.
Такой подход позволяет уже сейчас создавать искусственные нейронные сети и строить кибернетические модели информационных процессов в мозге более сложных животных вплоть до человека. Учитывая то, что экспериме6нты на мозге людей нельзя проводить по морально-этическим соображениям, путь создания кибернетических моделей с целью получения экспериментальных сведений о структуре и функциях человеческого мозга представляется весьма перспективным. Конечной целью при этом могут служить не только новые сведения о мозге, о механизмах его самооптимизации, самоорганизации, самопрограммирования, но и новые идеи, необходимые для построения нетрадиционных мозгоподобных суперЭВМ – нейрокомпьютеров.
2.Математическая модель информационных процессов в нейроне
|
где P(t) – мембранный потенциал участка аксона; H – локальный сдвиг мембранного потенциала за счет поступления внешнего воздействия; C – удельная емкость мембраны; q[P(t)] – проводимость мембраны участка аксона.
Экспериментально установлено, что если мембранный потенциал P(t) остается ниже некоторой пороговой величины QП, то проводимость мембраны практически не меняется. Однако в те моменты времени, когда под влиянием внешних раздражителей H потенциал P(t) достигает величины порога QП, электрически управляемые белки открывают свои шлюзы, что приводит к резкому изменению проводимости мембран. Аналитически данное обстоятельство можно выразить следующим образом:
|
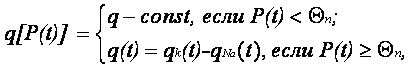
где qK(t) – переменная проводимость мембраны по отношению к ионам калия; qNa(t) – переменная проводимость мембраны по отношению к ионам натрия.
Однако ввиду того, что аксон является составной частью клетки, а его мембрана является клеточной мембраной, вполне убедительно мнение о возможности распространении этой теории на нейрон в целом, и в частности, на аксонный холмик, где осуществляется запуск потенциала действия.
При таком подходе в качестве правой части уравнения (1) можно использовать не только локальный сдвиг мембранного потенциала H, но и внутриклеточный потенциал V(t), формируемый за счет синаптических процессов в дендритном дереве и соме нейрона. Наиболее распространенная математическая модель данного процесса представляет собой алгебраическую сумму произведений всех входных потенциалов действия на соответствующие синаптические веса:
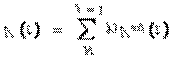 | |||
| |||