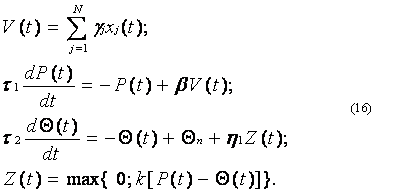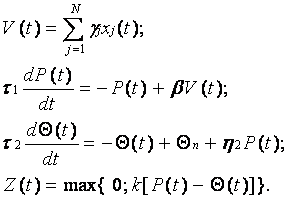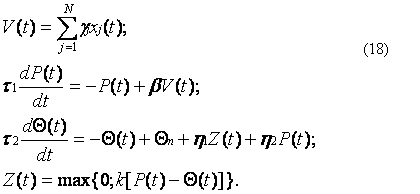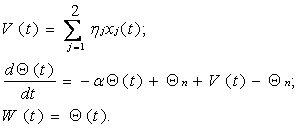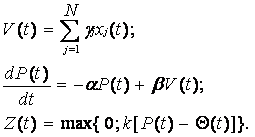Однако, если в качестве выходной величины неадаптивной модели нервной клетки использовать не функцию Z(t), а абсолютные значения дифференциалов dy(t), то выходная реакция такой модели на входное ступенчатое воздействие будет описываться соотношением
Нетрудно видеть, что график функции (14) качественно не отличается от переходной характеристики модели, реализующей “on” и “off” ответы нейронов. Последнее обстоятельство позволяет утверждать, что для моделирования простейших адаптивных реакций нервных клеток рецепторного типа достаточно воспроизводить первые два уравнения модели (12), а выходы организовать в соответствии с получаемым из (12) соотношением
где Q – неизменный во времени порог.
В то же время, по мнению нейрофизиологов, более сложные механизмы адаптивного поведения нейронов основаны на изменении их пороговых свойств. В соответствии с другими представлениями изменение порога клетки происходит в зависимости от изменения входной активности нейрона. Модель, воспроизводящую второй механизм, называют адаптивной по входу. Очевидно, что могут иметь место динамические нейроподобные элементы, адаптивные как по входу, так и по выходу одновременно. Строятся такие модели на основе следующих рассуждений.
Для построения на основе алгоритма (10) математической модели адаптивной обработки информации в нейроне будем исходить из того, что, по мнению физиологов, механизм адаптивного поведения нервных клеток связан с изменениями порога Q. В связи с этим используем уравнение (6), но не для формирования спайков как в модели (8), а для воспроизведения адаптивных реакций нейрона. При этом будем полагать, что при адаптации по выходу мембранный потенциал клетки сравнивается с переменным порогом Q(t), закон изменения которого имеет вид
Используя уравнение (15) в алгоритме (10), получим нейроподобную модель с адаптацией по выходу:
В случае адаптации по входу в правой части уравнения (15) вместо Z(t) необходимо использовать функцию P(t). Поэтому нейроподобная модель с адаптацией по входу несколько отличается от модели (16) и имеет следующий вид:
Очевидно, что для модели с адаптацией как по входу, так и по выходу будем иметь:
В системе уравнений (18) порог Q(t) зависит как от входной величины h2P(t), так и от выходной h1Z(t) активности нервной клетки. Причем полагая h2 = 0, h1 ¹ 0, получим модель с адаптацией только по входу. Более того, при h1 = h2 = t2 = 0 и R(t)–Q = y(t) получим исходную неадаптивную модель (10).
Иными словами, модель (18) более универсальна, чем неадаптивная модель (10), и по этой причине может быть использована для построения искусственных нейронов, воспроизводящих как адаптивные, так и неадаптивные реакции. Однако структура искусственного нейрона при этом также усложняется. Поэтому прежде чем решить вопрос о целесообразности подобного усложнения, необходимо учесть мнение физиологов о том, что нервная клетка является лишь структурной единицей мозга и в полной мере обладает далеко не всеми свойствами биологических систем.
В частности, многие физиологи полагают, что адаптивные реакции типа привыкания, как и многие другие функции нервной ткани, реализуются не отдельными нейронами, а их совокупностями в процессе совместной корпоративной деятельности. В связи с этим наряду с понятием нейрона, как структурной единицы нервной системы, в современной нейрофизиологии используется понятие о ее функциональной единице, в качестве которой выступает не отдельный нейрон, а некоторая совокупность нервных клеток, называемая нейронным ансамблем. В простейшем случае нейронный ансамбль состоит из двух взаимосвязанных нейронов, один из которых выполняет основные функции, а второй – вспомогательные (усиление, торможение, модификация процессов в основном нейроне и т. д.).
При таком подходе систему уравнений (18) можно рассматривать как модель информационных процессов не в отдельной клетке, а в гипотетическом двухнейронном ансамбле, основной нейрон которого реализует алгоритм (10), а вспомогательный воспроизводит процесс модификации порогового потенциала основного нейрона в функции от его входной и выходной активности. Алгоритм вспомогательного нейрона при этом может быть представлен в следующем виде:
где x1 = P(t) – пространственный потенциал дендритного дерева основного нейрона, поступающий на вспомогательный нейрон при помощи дендритных связей; x2 = Z(t) – выходная активность основного нейрона, заводимая на вспомогательный нейрон при помощи аксосоматической связи; Qп – порог покоя вспомогательного нейрона, совпадающий с порогом покоя нейрона основного; W(t) – соматический выход вспомогательного нейрона, поступающий на сому основного нейрона через сомасоматический контакт.
С учетом алгоритма (19), модель информационных процессов в основном нейроне принимает вид: